Carnet Botanique
Les fougères méditerranéennes du massif de la Clape
Survivre entre ombre et sécheresse
Explorations et photographies de AC et Ma Clape
Carnet Botanique
Survivre entre ombre et sécheresse
Explorations et photographies de AC et Ma Clape

Introduction
Juillet dans le massif de la Clape. Le calcaire renvoie la lumière, le Cers a séché la garrigue. Sur les plateaux, le ciste cotonneux, le romarin et le lentisque tiennent, feuilles dures, tiges grises. Au pied d’une paroi nord, dans une fissure ombragée, il y a une touffe rétractée, couverte d’écailles argentées, d’aspect parcheminé. Sous les feuilles sèches accumulées, c’est du vert qui va réapparaître à la prochaine pluie.
✦ ✦ ✦
Ce sont des plantes vasculaires: elles ont de vrais vaisseaux conducteurs de sève, comme les arbres ou les herbes, mais sans fleurs ni graines. Elles se reproduisent par des spores, des cellules infimes et légères ; ces spores minuscules (34 à 75 µm selon les espèces décrites dans ce carnet, voir les tailles par espèce) peuvent voyager sur de longues distances par anémochorie. Une spore passe là où aucune graine n'entre: une fissure de quelques millimètres sur une paroi verticale, l'interstice entre deux pierres. C'est leur grande liberté de dispersion, et la raison pour laquelle on les trouve dans des endroits aussi improbables que les falaises calcaires de la Clape.
Dans notre imaginaire collectif, les fougères évoquent l’ombre humide des sous-bois, le bord des ruisseaux. Le massif leur oppose l’inverse : un karst calcaire sec, exposé, qui ne retient pas l’eau. Ces fougères n’ont pas trouvé un refuge par hasard : elles ont réussi à s’implanter dans un milieu qui semble fait pour les exclure. C’est cela qu’il s’agit de comprendre.
Ce massif karstique se distingue par une perméabilité élevée: la structure fissurée de la roche favorise une infiltration rapide, limitant le ruissellement de surface en dehors des épisodes pluvieux intenses et des zones de lapiaz nu.
Et pourtant, trois espèces persistent. Elles ne sont ni rares ni spectaculaires au premier regard, mais chacune privilégie une réponse dominante face au stress hydrique. Ces réponses peuvent se combiner ; c'est la dominante de chaque espèce qui est mise en avant ici.
Ce carnet n'est pas un inventaire. C'est une enquête sur trois façons d'être au monde dans un endroit qui ne vous a pas attendu.
Le parcours du carnet
Portrait des fougères
Reconnaître les trois espèces sur le terrain: morphologie, saisons, habitats dans la Clape.
Le cadre environnemental
Comprendre les contraintes physiques du milieu et le rôle des communautés pionnières.
Les stratégies de survie
Entrer dans le détail des trois espèces: résurrection, plasticité phénologique, évasion spatiale.
Le cycle biologique
Examiner comment ces espèces bouclent leur cycle dans un milieu qui leur laisse peu de marges.
Lecture express · 5 minutes
Pour qui n'a pas le temps des quatre chapitres, la trame courte :
Pour aller plus loin : portraits sur le terrain · cadre environnemental · stratégies détaillées · cycle reproductif.
Dans ce carnet, le mois affiché sous chaque photographie est celui de la prise de vue. Pour ces trois espèces, la date compte: une même fronde a un aspect très différent selon la saison. Les saisons suivent la convention météorologique : printemps à partir du 1er mars, été à partir du 1er juin, automne à partir du 1er septembre, hiver à partir du 1er décembre.
Dans le massif, trois espèces de fougères se retrouvent régulièrement. Les reconnaître sur le terrain, c'est le point de départ de tout le reste: chaque espèce a une morphologie propre, une saison d'activité et un habitat qui lui correspondent. Ces trois paramètres sont aussi les premières clés pour comprendre pourquoi chacune a développé une stratégie différente face à la sécheresse.
Les trois espèces ont chacune un caractère immédiatement visible sur le terrain. Pour le Cétérach, ce sont les écailles rousses et denses qui tapissent la face inférieure des frondes, visibles à l'œil nu, diagnostiques à distance. Pour le Polypode, c'est la saison : en plein hiver, il déploie de grandes frondes vert vif avec des sores orange, le seul des trois à arborer le morphotype de fougère typique à cette saison. Pour la Doradille, c'est le rachis brun foncé et luisant, persistant longtemps après la chute des pinnules. Ces trois indices suffisent dans la grande majorité des cas.

Fronde allongée, découpée en lobes arrondis non séparés jusqu'au rachis ( pennatifide ). Longueur habituelle: 5 à 20 cm. Face supérieure vert sombre, face inférieure entièrement masquée par un feutrage dense d'écailles rousses à argentées, les paléas . En été, les frondes se roulent sur elles-mêmes, exposant ce feutrage argenté vers l'extérieur.
Toucher: la face inférieure est sèche et légèrement rugueuse, comme du papier de verre fin. En période active, les frondes sont souples et flexibles. En été, après dessiccation, elles sont friables: on les croirait mortes, et on les briserait d'une pression. Ce contraste saisonnier entre la souplesse hivernale et la fragilité estivale est l'indication la plus directe de l'anhydrobiose, avant même de regarder la couleur.
Nervation: les nervures secondaires forment des anastomoses marginales, caractère propre au sous-genre Ceterach , absent chez A. trichomanes , dont les nervures sont libres, comme chez la majorité des Asplenium Van den Heede et al., 2003, fig. 1 . Ce trait, combiné à l'absence d' indusie visible, a longtemps suffi à maintenir le Cétérach dans un genre distinct (voir encart Classification ci-dessous) avant son rattachement aux Asplenium par les phylogénies moléculaires.
Habitat: parois et fissures calcaires pouvant être exposées au soleil, lapiaz . C'est la seule des trois espèces à coloniser des stations vraiment ensoleillées.
 Hiver
Hiver
Asplenium ceterach touffe adulte Janvier
 Printemps
Printemps
Asplenium ceterach début d'enroulement · Avril
 Hiver
1 / 2
Hiver
1 / 2
Les écailles clathrées (réseau translucide argenté) masquent les sores à l'œil nu ; les sores restent fonctionnels sous les paléas. On distingue ici plusieurs amas de sporanges à différents stades: fermés (brun-noir) et partiellement ouverts (dorés).
 Printemps
1 / 1
Printemps
1 / 1
Plante en croissance: Une jeune fronde se déroule en crosse, entièrement enveloppée de paléas argentées. Les frondes adultes de la même touffe sont déjà étalées et actives. En bas, la face inférieure roux-brun annonce les sores en cours de maturation. Trois stades coexistent sur le même individu.
Le Cétérach peut s'accommoder de parois plus exposées, y compris en plein soleil.
L'espèce suivante fait le choix inverse: elle évite entièrement l'été et les stations ensoleillées, et ne se montre qu'à partir de l'automne, sur les versants que le soleil n'atteint pas.

Fronde vert clair à vert moyen, pennatiséquée : les segments (pennes) sont bien séparés jusqu'au rachis central, à bord légèrement dentelé. Longueur: 10 à 40 cm. Face inférieure portant des sores ronds, non recouverts d'indusie, disposés de part et d'autre de la nervure médiane de chaque penne, facilement visibles de novembre à mars.
Toucher: la fronde est coriace et un peu épaisse sous le doigt, sans la souplesse d'une feuille ordinaire. En hiver, elle reste ferme même par temps humide: aucune tendance à coller ou à s'affaisser. Les sores ronds sur la face inférieure, quand ils sont mûrs, laissent une légère trace orangée sur le doigt. C'est le moyen le plus simple de vérifier que la sporulation est en cours.
Habitat: parois et blocs rocheux calcaires orientés au nord ou nord-ouest. Bases de falaises ombragées. Parfois épiphyte sur les branches basses d'arbres en milieu humide.
 Hiver
Hiver
Sores à maturité · Janvier
 Printemps
Printemps
Rhizome exposé · Mars
Là où le Polypode joue sur la saison pour éviter la sécheresse, la Doradille joue sur l'espace. Elle recherche les parois orientées nord et les fissures ombragées, là où les quelques centimètres de roche qui la séparent de l'extérieur suffisent à changer les conditions.

Fronde étroite, longue de 5 à 20 cm, portant de petites pinnules arrondies ( pennes ) de part et d'autre d'un rachis central brun foncé, brillant, très persistant. C'est ce rachis sombre et luisant qui distingue immédiatement l'espèce. Les pinnules ne tombent pas lors de la sécheresse estivale : elles la traversent en maintenant leur activité au minimum, puis chutent par sénescence progressive après plusieurs saisons. Le rachis, lui, persiste bien au-delà : visible mois après mois, parfois des années après la perte des pinnules. Sores allongés ( indusiés ), disposés sur la face inférieure des pinnules, distincts au début de la sporulation puis devenant confluents à maturité jusqu'à recouvrir presque toute la face inférieure de la penne Page, 1997 .
Toucher: le rachis est le critère tactile le plus net de l'espèce: lisse, rigide, légèrement froid au doigt, il se distingue immédiatement des rachis verts et plus souples des deux autres fougères. Les pinnules, quand elles sont présentes, s'écrasent facilement sous l'ongle. Le rachis, lui, résiste. Cette différence de rigidité entre les deux parties de la même fronde dit quelque chose sur leur durée de vie respective.
Habitat: fissures étroites de rochers et de vieux murs calcaires, ombragées ou mi-ombragées. Espèce strictement chasmophyte .
 Hiver
Hiver
Asplenium trichomanes, rachis brun caractéristique · Février
 Printemps
Printemps
Asplenium trichomanes, rachis et sores · Mars
 Printemps
Printemps
Asplenium trichomanes, rachis et sores · Mai
 Printemps
Printemps
Asplenium trichomanes , rachis et pinnules · Mai
Les trois critères les plus utiles sur le terrain: la face inférieure de la fronde, le rachis et la saison d'observation. Le tableau ci-dessous résume les caractères diagnostiques essentiels.
Tableau morphologique comparatif
Critères de détermination sur le terrain
| Critère |
Cétérach Asplenium ceterach |
Polypode austral Polypodium cambricum |
Doradille capillaire Asplenium trichomanes |
|---|---|---|---|
| Taille | 5–20 cm | 10–40 cm | 5–20 cm |
| Découpage du limbe | Pennatifide: lobes arrondis non séparés jusqu'au rachis | Pennatiséqué: pennes bien séparées jusqu'au rachis | Pennatiséqué: petites pinnules arrondies régulières |
| Rachis | Vert, couvert d'écailles rousses | Vert à brun clair, peu visible | Brun foncé, brillant : critère immédiat |
| Face inférieure | Feutrage dense d'écailles rousses à argentées : critère immédiat | Verte, sores ronds bien visibles | Verte, sores allongés sur chaque pinnule |
| Sores | Masqués par les écailles: invisibles à l'œil nu | Ronds, nus (sans indusie), jaunes à orangés en hiver | Allongés, indusiés, disposés obliquement |
| Indusie | non ou vestigiale (masquée par les paléas) | Absente | Présente, linéaire |
| Saison d'observation |
Automne–printemps Brunâtre et roulée en été |
Automne–hiver–printemps Absente en été |
Toute l'année Ralentie en été, rachis toujours visible |
| Exposition | Toutes expositions, y compris plein soleil | Nord, nord-ouest: ombre obligatoire | Mi-ombre à ombre: fissures profondes |
| Critère décisif | Face inférieure argentée–rousse, visible même sur plante desséchée | Sores ronds et nus sur fond vert clair, présents uniquement en hiver | Rachis brun brillant persistant, visible après chute des pinnules |
Ce qu'il faut retenir
Les trois espèces ont des exigences distinctes: le Cétérach sur les parois exposées, le Polypode sur les versants à ombre hivernale, la Doradille dans les fissures ombragées. Mais ce ne sont pas des territoires: dans une fissure assez profonde, Cétérach et Doradille coexistent, séparés de quelques centimètres par un gradient d'humidité et de lumière. Pour comprendre comment ces lignées en sont arrivées là, il faut revenir à leur histoire.
Une précision sur le titre de ce carnet: les trois espèces appartiennent toutes à l'ordre des Polypodiales , un rang taxonomique qui englobe à la fois les Aspleniaceae ( Asplenium ceterach , A. trichomanes ) et les Polypodiaceae ( Polypodium cambricum ). C'est ce cadre commun, et non la seule famille des Polypodiaceae, qui justifie le regroupement sous ce nom. La section suivante revient sur l'histoire évolutive de cet ordre.
Ces trois espèces ne sont pas arrivées dans les fissures de la Clape par hasard. Elles appartiennent à une lignée dont les premières traces remontent au Carbonifère, il y a 360 millions d'années, bien avant les plantes à fleurs, bien avant les dinosaures. Comprendre d'où elles viennent aide à comprendre pourquoi elles sont encore là.
Les fougères que l'on connaît aujourd'hui ne sont pas des rescapées directes du Carbonifère. Les espèces des genres Asplenium et Polypodium appartiennent aux Polypodiales , un ordre dont la lignée remonte au Trias (~220 Ma) et dont la diversification principale se situe au Jurassique inférieur à moyen (180-160 Ma) Schuettpelz & Pryer, 2009 . Les estimations restent débattues selon les calibrations moléculaires utilisées Du et al., 2021 ; Nitta et al., 2022 , sans consensus établi, mais convergent sur une apparition au Mésozoïque , bien après l'effondrement des grandes forêts carbonifères. Ce sont des fougères dites « modernes », issues d'une radiation évolutive récente à l'échelle géologique. Après l'extinction K-Pg, il y a 66 millions d'années, les fougères ont été parmi les premières plantes à recoloniser les paysages dévastés par l'impact d'astéroïde, un épisode documenté dans les sédiments sous le nom de Fern Spike . Les trois espèces de la Clape descendent de lignées qui ont traversé cet événement.
Le plan d'organisation des fougères de ce carnet (fronde divisée, rhizome rampant, reproduction par spores) est établi depuis le Jurassique et les archives fossiles n'attestent aucune modification majeure de leur architecture depuis lors ( Schneider et al., 2004 ; Schuettpelz & Pryer, 2009 ). Quelque chose a fonctionné d'emblée et n'a pas eu besoin d'être modifié.
Cette histoire longue a laissé des traces dans les noms que portent ces plantes. Fougère vient du latin populaire, par l'évolution phonétique ordinaire du roman. Mais les noms des genres Asplenium et Polypodium viennent du grec médical antique, et celui du Cétérach arrive par la médecine arabe médiévale. Trois plantes, trois trajectoires de noms, qui reflètent les différentes couches de savoirs botaniques dont la nomenclature européenne est faite.
Étymologie, Fougère
Du latin populaire au français courant
Le mot
fougère
est attesté en vieux français sous la forme
fougiere
dès le XII
e
siècle. On le retrouve au XVI
e
siècle sous cette forme
fougiere
mais aussi
feuchiere
(Histoire générale des plantes, Dalechamps, Jacques 1513-1588).
Il descend du bas latin
*filicaria
, qui signifiait proprement « fougeraie » (un lieu où poussent les fougères en abondance), le français a hérité du nom du lieu, pas directement du nom de la plante.
*Filicaria
est lui-même dérivé du latin classique
filix, filicis
, qui désignait la fougère dans l'Antiquité romaine (Virgile, Pline l'Ancien & Columelle). Elle était souvent associée à l'adjectif
infelix
chez ces auteurs latins, non pas parce qu'elle porte malheur, mais parce qu'elle est « inféconde » car elle ne produit pas de fruit comestible.
Pline l'Ancien note dans son traité des plantes que les fougères « n'ont ni fleurs ni graines » (
neque flos iis ullus neque semen
), observation exacte que la botanique moderne n'a fait que préciser
Pline l'Ancien,
Naturalis Historia
, XXVII, § LV
.
La voie populaire
filix → *filicaria → fougière → fougère
L'astérisque (*) signale une forme reconstruite: non attestée dans les textes, mais déduite par comparaison linguistique.
C'est la trajectoire du mot tel qu'il s'est transmis oralement, de génération en génération, sans passer par l'écrit savant. Le résultat est le mot de tous les jours, celui qu'on emploie sur le terrain sans y penser.
La voie savante
filix → filicula → filicaire, filicophyte
Au XVI e siècle, les botanistes humanistes (Fuchs, Mattioli, Dodoens) utilisent dans leurs manuscrits latins le diminutif filicula (« petite fougère » comme on a pu le voir sur l'illustration du manuscrit de Gerardo Cibo Ass MS 22332), forme attestée dans les textes, contrairement à *filicaria. Le même radical filix a ensuite donné, par emprunt direct à l'époque moderne, les termes techniques filicaire (qui concerne les fougères) et filicophyte (nom savant du groupe). On le retrouve dans Dryopteris filix-mas , la fougère mâle commune.
φύλλον
, la feuille) a parfois été proposé pour expliquer le latin
filix
, mais cette parenté reste discutée: les linguistes ne l'ont pas établie avec certitude. Ce qui ne change rien à l'usage
Les fougères avaient été positionnées dans un groupe appelé Ptéridophyte qui contenait aussi les Lycophytes : les lycopodes (le lycopode en massue, Lycopodium clavatum , commun en forêt européenne), les sélaginelles (la sélaginelle denticulée, Selaginella denticulata , sur rochers méditerranéens) et les isoètes (plantes aquatiques à feuilles en alène), une lignée apparue au Dévonien. Le regroupement était morphologique: toutes ces plantes ont des vaisseaux conducteurs, se reproduisent par spores, sans fleurs ni graines. Ce que la botanique du XIX e siècle ne voyait pas encore, c'est que ces ressemblances recouvrent une séparation vieille de 400 millions d'années. La phylogénie moléculaire a tranché: fougères et Lycophytes sont deux lignées distinctes, et les classifications modernes les traitent comme telles.
Étymologie, Ptéridophytes
Du grec ancien, deux racines combinées
Première racine
Pteris / Pteridos
πτερίς / πτερίδος
Vient de
pteron
(
πτερόν
), qui signifie
aile
ou
plume
. Les Grecs désignaient ainsi la fougère, dont les frondes découpées rappellent visuellement la forme d'une plume d'oiseau.
Deuxième racine
Phyton
φυτόν
Du verbe
phuein
(
φύειν
), qui signifie
faire pousser
,
naître
,
croître
. Il désigne simplement la plante ou le végétal.
Le terme fut codifié au XIX e siècle dans le cadre de la systématisation de la botanique scientifique. Le botaniste allemand Wilhelm Hofmeister contribua vers 1850 à clarifier le cycle de reproduction de ces plantes, un travail essentiel pour comprendre ce qui les distingue des plantes à fleurs.
Avant de fermer ce premier portrait, une question utile pour situer ces trois espèces parmi les fougères du Languedoc : pourquoi celles-ci, et pas les autres ?
Avant de continuer
Ces trois portraits posent le cadre: on sait maintenant reconnaître les espèces, on connaît leur saison et leur habitat préféré. Ce que l'on ne sait pas encore, c'est comment un massif aussi sec et perméable peut les retenir. La section suivante l'examine: les contraintes physiques du milieu, et les organismes qui, avant les fougères, ont préparé le terrain.
Le massif de la Clape impose des conditions extrêmes: un socle calcaire perméable, un ensoleillement brutal et presque aucune eau en surface. Les fougères ne s'y installent pas par hasard. Pour comprendre pourquoi elles sont là où elles sont, il faut d'abord comprendre la roche elle-même: sa nature détermine la chimie, la structure détermine l'eau.
Le massif de la Clape impose deux contraintes physiques majeures aux plantes qui s’y installent: une roche calcaire qui contrôle la chimie des nutriments, et une hydrologie qui ne laisse presque aucune eau en surface.
Le massif est constitué de plusieurs unités sédimentaires du Crétacé inférieur empilées, dont une partie en faciès urgonien (calcaires blancs compacts à rudistes, polypiers et bryozoaires). Les stations enregistrées dans ce carnet se situent principalement sur les
Calcaires moyens du Plan de Roque
(Aptien supérieur, environ 40 m d'épaisseur), qui forment les entablements des plateaux du Plan de Roque et des Auzils
Fauré, 2022
, sans s'y limiter, d'autres bancs urgoniens du massif portant également des stations.
Cette roche, dure, blanche, extrêmement fissurée, induit une perméabilité verticale quasi totale. Les précipitations, parfois violentes lors des épisodes méditerranéens, ne ruissellent que brièvement en surface avant de s'infiltrer dans les calcaires. Une unité marneuse d'une quatre-vingtaine de mètres d'épaisseur compartimente le massif : les calcaires urgoniens supérieurs, où se trouvent les stations de fougères, déchargent leurs eaux en sources temporaires à leur contact avec cette couche semi-imperméable
Khaska et al., 2013
.
Sur les plateaux et les falaises, le sol se résume à des
lithosols
squelettiques logés dans les fissures du lapiaz : quelques centimètres d'épaisseur au mieux, soumis à des fluctuations thermiques et hydriques extrêmes. Ce substrat ne peut accueillir que des espèces capables de tolérer ou d'éviter la sécheresse, les deux stratégies examinées dans la partie suivante.
À cela s'ajoute un verrou nutritif propre aux milieux carbonatés, détaillé dans le chapitre suivant.

Régime pluviométrique, massif de la Clape
Station Météo-France n° 11262005 · Narbonne · 110 m · normale 1991-2020 · série 1989-2024
La station Météo-France de Narbonne enregistre 635 mm en moyenne sur la période 1991-2020, valeur retenue comme normale climatologique de référence. Sur la décennie récente 2015-2024, la moyenne descend à 509 mm, soit un recul d'environ 20 %. L'amplitude interannuelle reste considérable : 328 mm en 2023, année la plus sèche de la série, jusqu'à 1205 mm en 1996. Un rapport de un à quatre d'une année à l'autre.
L'été est structurellement sec. Juillet et août réunis totalisent 48 mm en moyenne. Le creux est centré sur juillet, mois le plus pauvre de l'année avec 17 mm en moyenne, parfois moins de 5 mm certaines années. Cette aridité n'est pas une anomalie. C'est la constante du régime méditerranéen, et les fougères du massif l'ont intégrée depuis longtemps dans leur calendrier.
L'essentiel des apports tombe en automne, sous forme d'épisodes courts. Octobre concentre 92 mm en moyenne, novembre 78 mm, septembre 61 mm. Au printemps, avril reste un mois pluvieux secondaire avec 64 mm.
Sur le karst de la Clape, ces apports s'infiltrent rapidement dans les calcaires fissurés. Une unité marneuse semi-imperméable bloque la percolation vers le bas : l'eau des calcaires urgoniens supérieurs ressort en sources temporaires à leur base, à la faveur de ce contact. L'eau est là, puis disparaît.
Cumul annuel · 2011-2025 (station 11262005)
Épisodes méditerranéens intenses : durée, total épisode et maximum journalier
L'épisode du 26 septembre 1992 (290 mm en une journée, 333 mm sur le mois) appartient à la séquence cévenole qui a frappé Vaison-la-Romaine quatre jours plus tôt. C'est le plus gros événement 24 heures enregistré à Narbonne sur la série 1950-2024. Le record absolu mensuel reste octobre 1965 avec 625 mm sur la longue série composite. Le cumul 24 heures mesure le pic d'intensité, pas la durée totale de l'épisode. Pour les fougères, c'est cette durée, et le film d'eau résiduel qu'elle entretient, qui conditionne la reproduction.
Ce que ces données signifient pour les fougères
La sécheresse estivale n'est pas une crise. C'est un paramètre fixe du climat narbonnais, prévisible à quelques semaines près, et les trois espèces l'ont intégrée dans leur calendrier. Le Cétérach entre en anhydrobiose avant l'épuisement de ses réserves. Le Polypode est dormant dès juin. La Doradille tient dans une fissure où quelques millimètres de roche décalent le bilan hydrique local.
Ce qui varie d'une année à l'autre, c'est l'automne. La fécondation des fougères exige un film d'eau liquide, durable : l' anthérozoïde multiflagellé nage jusqu'à l'archégone. Sans humidité au sol sur plusieurs jours consécutifs, la reproduction échoue. Sur la longue série Narbonne 1950-2024, un automne sur cinq cumule moins de 80 mm en septembre-novembre, seuil sous lequel la fenêtre de fécondation reste à peine entrouverte. Cette variabilité est ancienne dans le climat local : les décennies 1950 à 1970 ont compté autant d'automnes secs que la période récente. Le détail figure dans Les fenêtres de reproduction .
Les épisodes intenses ne changent pas ce bilan. Cent ou deux cents millimètres en 24 heures remplissent les fissures, puis le karst absorbe. Ce qui compte pour le prothalle d'une fougère, c'est la durée du film d'eau résiduel, pas le volume tombé. Sur ce point, le signal d'inquiétude vient moins de la pluviométrie elle-même que de la demande évaporatoire, traitée dans l'encart suivant.
Bilan hydrique : la pluie ne dit pas tout
Station Météo-France n° 11262005 · ETP Penman-Monteith · 1989-2024
L'évapotranspiration potentielle (ETP) mesure la quantité d'eau qu'une surface de référence standardisée (gazon court, bien irrigué) restituerait à l'atmosphère par évaporation et transpiration, dans les conditions de température, d'humidité, de rayonnement et de vent observées. Cette demande atmosphérique est indépendante de la végétation réelle du site.
À Narbonne, elle atteint 1241 mm par an en moyenne (méthode Penman-Monteith), soit le double des précipitations.
En comparaison, l’évapotranspiration réelle (ETR) est limitée par la disponibilité en eau. Sur la période 1991-2020, avec une pluviométrie moyenne de 635 mm/an, l'ETR est estimée à environ 400 mm/an (selon la répartition 60/35/5 du karst méditerranéen, Bakalowicz, 2005). Ce plafond varie fortement selon les précipitations annuelles, passant d'environ 300 mm lors de l'année sèche de 2024 (466 mm de pluie) à plus de 780 mm en 1996 (1 205 mm de pluie).
Le déficit hydrique réel, soit l'écart entre l'ETP et l'ETR, atteint environ 840 mm. Le bilan précipitations moins ETP donne le déficit hydrique potentiel : −606 mm par an (La région perd plus d'eau qu'elle n'en reçoit). Ce déséquilibre explique pourquoi le film d'eau résiduel sur le lapiaz s'évapore en quelques heures après un épisode pluvieux : le pouvoir évaporant de l'air surpasse largement l'eau réellement disponible, un phénomène accentué par la nature du karst de la Clape.
En deux décennies, ETP et pluie évoluent en sens opposés. Entre la période 1989-2004 et la période 2009-2024, la demande évaporatoire annuelle a gagné 92 mm et le déficit hydrique s'est creusé de −502 à −750 mm par an. Saison par saison, la bascule est nette : l'hiver passe d'un bilan moyen excédentaire de +63 mm à un bilan déficitaire de −31 mm, l'automne du même mouvement (+36 mm vers −51 mm). Ces deux saisons sont précisément celles où le Cétérach germe à l'automne et où le Polypode est actif en hiver. La demande évaporatoire dépend de la température, du rayonnement et du vent : le Cers, sec et fréquent (170 à 250 jours estimés par an), est l'un des moteurs de cette hausse ; il accélère l'évaporation à la surface des rochers et des fissures indépendamment de la saison.
Conséquence pour les fougères
Sur la même quantité de pluie, l'eau utile diminue. Le film d'eau résiduel après un épisode automnal sèche plus vite. La fenêtre de fécondation se rétrécit. La dormance estivale doit être déclenchée plus tôt, sa sortie attend plus longtemps. L'aridification ne se mesure pas seulement en millimètres tombés du ciel, elle se mesure aussi en millimètres prélevés par l'air.
L'eau de pluie ne reste guère en surface. Elle se charge en CO₂ au contact de la matière organique du sol (respiration microbienne et racinaire), puis s'infiltre dans les fissures du calcaire, qu'elle dissout lentement au fil de sa descente.
Ce processus, répété depuis des millénaires, a creusé un réseau de fissures et de galeries souterraines dans lequel s'infiltre l'eau qui n'est ni évaporée ni transpirée par la végétation. Sur un karst méditerranéen comme la Clape, le bilan hydrique annuel moyen se répartit en trois postes: l' évapotranspiration prélève environ 60 % des précipitations, l'infiltration souterraine en mobilise environ 35 %, se répartissant entre l'aquifère supérieur des calcaires urgoniens et l'aquifère profond selon une proportion non quantifiée pour la Clape Khaska et al., 2013 , et il ne reste qu'une fraction de l'ordre de quelques pour cent retenue localement sur les substrats rocheux par capillarité et condensation, ordre de grandeur estimé par différence sur les bilans de karsts méditerranéens de basse altitude Bakalowicz, 2005 ; Mangin, 1975 , sans mesure directe pour la Clape. Un épisode méditerranéen violent peut saturer la surface quelques heures: dès qu'il cesse, le lapiaz est sec. Pas de cours d'eau permanent, pas de nappe accessible depuis la surface. À la base des calcaires urgoniens, quelques résurgences temporaires rejaillissent après les fortes pluies et trahissent le transit souterrain, sans alimenter aucune zone humide durable dans le massif lui-même.
Ce que retient le massif est localisé et discret. Dans les fissures les plus étroites, la tension de surface maintient un film d'eau contre la roche plusieurs jours après la dernière pluie. Sur les parois orientées au nord, les températures plus basses favorisent la condensation nocturne: l'air humide de la nuit dépose un film d'eau sur la roche froide. Ces deux phénomènes sont minimes. Ils suffisent aux organismes dont les besoins sont ajustés à ce niveau. Les lichens arrivent en premier, les mousses dans leurs traces, et les fougères là où le substrat commence à ressembler à quelque chose.
Les flux de l'eau dans le massif calcaire
De ces trois flux, seul le dernier est directement accessible aux plantes. Les 60 % d'évapotranspiration correspondent à un bilan annuel moyen : lors des épisodes intenses, l'eau disparaît presque entièrement dans les calcaires sans s'évaporer, comme le décrit la section précédente. Entre les épisodes, sur le lapiaz nu, c'est l'évaporation directe depuis la roche qui domine, la transpiration des plantes y étant marginale.
L'eau utile est celle que retient l' épikarst par capillarité dans ses micro-fissures. C'est précisément pendant et juste après les épisodes pluvieux que cette couche altérée superficielle du calcaire se charge, piégeant l'eau sans qu'elle s'évapore ni ne parte en profondeur. Dans les jours et semaines qui suivent, la fraction infiltrée plus bas ressort en sources temporaires à la base des calcaires urgoniens ou rejoint l'aquifère profond, tandis que l'épikarst restitue progressivement son eau aux racines, disponible plusieurs jours à plusieurs semaines Perrin et al., 2003 ; Aquilina et al., 2006 . C'est ce décalage temporel qui fait de l'épikarst le réservoir utile pour les fougères, là où l'infiltration souterraine n'est qu'une perte.
Cette absence de sol profond a une conséquence directe sur la nutrition des plantes. Mais il y a une deuxième contrainte, moins visible et tout aussi déterminante: le calcaire lui-même bloque chimiquement une partie des nutriments disponibles, avant même que les racines ne puissent les atteindre.
Comme toute plante vasculaire, les fougères ont besoin d'azote (synthèse des protéines et de la chlorophylle), de phosphore (transfert d'énergie, ADN, membranes), de potassium (régulation osmotique, ouverture des stomates), de calcium et de magnésium (structure cellulaire et centre de la chlorophylle), ainsi que d'oligoéléments dont le fer est le plus critique (transfert d'électrons dans la chaîne photosynthétique). Le phosphore et le fer sont bien présents dans la roche: le phosphore sous forme de traces d' apatites incorporées lors de la sédimentation crétacée, le fer sous forme d'oxydes disséminés dans la matrice calcaire et concentrés dans la terra rossa des fissures. Sur un substrat calcaire à pH 7,5–8,5, les deux deviennent quasi inaccessibles. C'est sur eux que se concentre le verrou nutritif de la Clape.
Le calcaire n'est pas un support neutre. En milieu carbonaté, le pH oscille entre 7,5 et 8,5, des valeurs qui bloquent une partie des nutriments avant même que les racines ne les atteignent. À ces valeurs, le phosphore précipite avec le calcium pour former des phosphates de moins en moins solubles: brushite d'abord, puis phosphate octacalcique, puis hydroxyapatite , une forme quasi inerte. Ce processus de rétrogradation est irréversible dans les conditions naturelles du massif. Parallèlement, le fer, le manganèse, le zinc et le bore deviennent indisponibles par précipitation ou adsorption sur les surfaces carbonatées.
Une roche chimiquement bien dotée peut donc nourrir très peu de chose. Les nutriments sont là, mais inaccessibles aux racines.
Le sol est donc doublement hostile: pauvre en eau accessible et pauvre en phosphore mobilisable. Mais le massif n'est pas homogène. Certains secteurs échappent partiellement à ces contraintes grâce à leur orientation et à leur géométrie. Les fougères s'y installent.
La Clape n'a plus de rivière. Quelques sources temporaires, des résurgences qui rejaillissent après les pluies, et c'est tout. Leur régime (intermittent, décalé de plusieurs jours après l'épisode pluvieux) est caractéristique des karsts méditerranéens de basse altitude à réseau de conduits bien développé Nicod, 2012 . Ce sont elles, pourtant, qui décident de la présence ou de l'absence des espèces hygrophiles .
Les falaises orientées nord jouent un rôle différent. Le Cers dessèche l'ensemble du massif sans distinction, mais ces parois reçoivent peu de soleil direct et se trouvent souvent dans l'ombre aérodynamique des reliefs qui les surplombent : le flux d'ONO bute sur la crête et accélère au-dessus, laissant la base de la paroi dans une zone de vitesse réduite. À l'inverse, les couloirs entre combes et défilés du massif concentrent et accélèrent le flux: l'effet Venturi y amplifie le dessèchement, mais s'accompagne d'un léger refroidissement par détente qui favorise la condensation nocturne sur les parois encaissées. C'est la combinaison de l'orientation et de cet abri partiel qui leur confère une humidité et une fraîcheur que le reste du massif ignore. À leurs pieds, les fougères s'y maintiennent, parce qu'elles échappent à la fois à la radiation directe et au plein effet desséchant du vent.
 Printemps
Printemps
Une des stations de fougères de la Clape · Avril
Au pied de cette falaise nord, les deux mécanismes décrits plus haut se superposent sur la même paroi: la capillarité dans les fissures de moins d'un millimètre retient un film d'eau plusieurs jours, la condensation nocturne en renouvelle chaque matin quelques dixièmes de millimètre. Une fissure de 0,5 mm peut conserver de l'humidité quand la roche exposée à un mètre de là sèche en quelques heures. Le volume est trivial ; ce qui compte pour un rhizome de fougère, c'est la régularité de l'apport. Cette régularité se concentre sur quelques mètres carrés à la base des parois nord, et c'est précisément là que les fougères du massif sont présentes.
Dans le massif, la ressource en eau suit un calendrier strict. Les pluies se concentrent à l'automne et au printemps. La sécheresse s'installe de juin à septembre, parfois dès mai lors des années sèches. Pour les fougères, ce calendrier définit les fenêtres où la fécondation est possible, où la germination d'une spore peut aboutir à un prothalle viable. Entre ces fenêtres, les fissures les plus profondes conservent une humidité résiduelle qui permet au rhizome de tenir, pas de se reproduire. Ce décalage entre disponibilité en eau et exigences du cycle reproductif est une contrainte que les adaptations morphologiques peuvent atténuer, jamais supprimer. Sa portée écologique se modifie avec la hausse de l'évapotranspiration : sur la même quantité de pluie, l'eau utile diminue. Le cycle biologique y revient précisément.
La fissure : point de convergence
Tout ce que cette section a décrit à l'échelle du massif se concentre dans un espace de quelques millimètres. La géologie urgonienne produit des fractures nettes à perméabilité quasi totale. L'hydrologie y dépose le seul film d'eau résiduel que le karst ne draine pas immédiatement. La chimie du calcaire y est localement modifiable par les racines, dans les premiers centimètres de profondeur. L'orientation de la paroi y détermine l'amplitude thermique et l'effet du Cers.
Une fissure n'est pas un sol. C'est une colonne verticale dont les conditions changent avec la profondeur : amplitude thermique maximale en surface, zone tampon à 3–5 cm, obscurité et humidité quasi constantes au-delà. Le rhizome des trois espèces occupe la zone tampon. Les frondes sortent en surface.
Trois paramètres distinguent les fissures habitables des fissures stériles : la largeur (seuil fonctionnel estimé autour de 5–8 mm, en dessous duquel l'accumulation organique est insuffisante), la profondeur (moins de 3 cm n'amortit pas l'amplitude thermique estivale), et l'orientation de la paroi. Ces valeurs sont des estimations par analogie avec des calcaires méditerranéens comparables ; aucune mesure directe n'a été publiée pour la Clape.
La fissure est aussi une unité de temps. Une fissure colonisée depuis plusieurs décennies conserve sa colonne organique d'une sécheresse à l'autre. Une fissure neuve, géométriquement identique, peut rester stérile pendant des décennies faute de biofilm initial. Ce que les organismes pionniers construisent dans la fissure ne se reconstitue pas rapidement.
L'aridité générale et ces refuges ponctuels expliquent en partie pourquoi chaque espèce s'est calée sur une niche différente ( Les stratégies de survie y revient en détail). Mais il reste une question préalable: comment les fougères s'installent-elles sur une roche nue, là où le sol n'existe pas encore? La réponse passe par ce qui les a précédées.
Regardez une falaise de la Clape: des traînées sombres sur le calcaire, une croûte grise par endroits, une touffe verte coincée dans une fissure. Ce désordre apparent est en réalité une chaîne, et chaque maillon prépare le suivant. Les fougères en sont le bout: elles ne s'installent pas sur la roche nue, mais sur un terrain que d'autres ont mis des décennies à rendre habitable.
Biofilm
La roche cesse d'être stérile: une pellicule de micro-organismes prend pied.
Lichens
La surface s'érode et se structure, les premières fissures se garnissent.
Mousses
Un premier matelas qui retient l'eau plusieurs jours après la pluie.
Succulentes
Les premières plantes à fleurs fixent un proto-sol où une spore peut germer.
La fougère ne germe pas sur la roche vraiment nue, mais la chaîne n'est pas un escalier obligé: tous les maillons ne sont pas toujours réunis. Les pages qui suivent les décrivent un à un.
Avant qu'un seul lichen puisse s'installer, la roche brute doit cesser d'être stérile. C'est le travail du biofilm, une pellicule quasi invisible de micro-organismes qui colonisent les premiers millimètres de la surface calcaire. On le voit sur les falaises de la Clape sous la forme de traînées sombres, grises ou noirâtres, qui n'ont rien à voir avec de la saleté: c'est de la vie microscopique organisée. Pensez à la patine qui verdit une pierre tombale ou un mur à l'ombre: même phénomène, une fine croûte vivante collée à la roche, sauf qu'ici elle travaille sur du calcaire en plein soleil.

Paroi calcaire sub-verticale : avant même les mousses et les lichens (stade dit précryptogamique )
Les cyanobactéries épilithiques , surtout les genres Gloeocapsa , Scytonema et Nostoc , agissent de deux façons. D'abord en surface: en adhérant à la roche et en y retenant l'humidité, le biofilm l'altère lentement et y piège des particules, ouvrant les premiers points d'ancrage. La dissolution proprement dite du calcaire, elle, vient surtout de l'eau de pluie chargée en CO₂, la dissolution karstique , et non du biofilm, qui ne sécrète pas d'acide. Ensuite côté azote: le Nostoc , reconnaissable après la pluie à ses amas gélatineux vert sombre dans les creux du lapiaz, fixe l'azote atmosphérique et le transforme en ammonium. À sa mort, il le libère sur la roche, un premier apport nutritif là où il n'y avait rien, dont profiteront les lichens qui suivent.
Attention cependant: toute cyanobactérie visible sur la roche n'est pas du biofilm. Le Nostoc dont il est question ici est libre, dans sa gaine et sans champignon associé. Le même genre peut aussi vivre enveloppé dans le thalle d'un champignon: on parle alors de cyanolichen , un organisme distinct, bien plus tardif, traité dans la section sur les lichens, plus bas.
 Hiver
1 / 3
Hiver
1 / 3
Surface calcaire couverte de Xanthoria parietina (lichen foliacé jaune-orangé). Deux jeunes frondes de Polypodium cambricum dans la fissure centrale: le lichen a préparé la surface, la fougère s'installe dans l'interstice qu'il ne peut pas occuper.
 Hiver
2 / 3
Hiver
2 / 3
Podéties en coupe à bords frangés caractéristiques. Ce Cladonia s'installe après les crustacés, dans les micro-accumulations organiques. Asplenium ceterach en bas: deux niveaux pionniers coexistent sur le même substrat.
 Hiver
3 / 3
Hiver
3 / 3
Colonie de Xanthoria parietina adulte en développement radial (plusieurs décennies). Sedum à feuilles lancéolées aplaties au premier plan: morphologie compatible avec Sedum gr. rupestre s.l., la seconde espèce documentée dans le massif. S. sediforme reste l'orpin dominant du lapiaz.
La roche calcaire nue est colonisée en premier par les cyanobactéries et les lichens . Ces organismes préparent lentement la surface de la roche (micro-altération, rétention de poussières), créant une microrugosité qui retient les particules et l'eau. Ils fixent l'azote atmosphérique et commencent à accumuler une fraction organique. Quelques millimètres par siècle, mais c'est suffisant pour ce qui suit.
Ce travail de surface est nécessaire mais insuffisant. Les lichens créent les conditions d'une colonisation, ils ne la complètent pas. C'est l'étape suivante, les mousses, qui transforme une roche légèrement érodée en un substrat où une fougère peut germer.
Les bryophytes , mousses et hépatiques, s'installent après les lichens. Une touffe de mousse dans une fissure fonctionne comme un réservoir: elle absorbe l'eau de pluie et la relâche sur plusieurs jours, long après que la surface sèche est à nouveau brûlante. Elle amortit aussi les écarts thermiques diurnes, qui peuvent dépasser 30 °C en surface rocheuse en plein été.
Ce phénomène est central dans la théorie de la facilitation : la mousse n'est pas une concurrente de la fougère, elle en est la condition préalable. Sans ce précurseur, le prothalle de la fougère, cette minuscule structure intermédiaire de quelques millimètres qui précède la vraie plante, ne survivrait pas aux premières semaines après la germination.
Une fois le coussin de mousse établi, un troisième type de plante entre en scène. Ni lichens ni fougères, les succulentes rupestres jouent un rôle de consolidation que les mousses seules ne peuvent pas assurer.
 Hiver
1 / 4
Hiver
1 / 4
Installation dans les fissures
Tiges stolonifères ligneuses courant sur le calcaire.
Les trous et la texture (Micro-karstification)
La surface de la roche montre une multitude de petites anfractuosités, de pores et de perforations. Ce faciès "troué" est le résultat d'une double action:
L'érosion chimique (dissolution) : L'eau de pluie, légèrement acide, dissout le carbonate de calcium, élargissant les micro-fissures.
L'action biologique (bio-érosion) : Traces de lichens crustacés grisâtres qui attaquent la pierre en surface, créant ce micro-relief idéal.
 Hiver
2 / 4
Hiver
2 / 4
Sedum sediforme (tiges rampantes), coussin de mousse (centre), jeune fronde de Polypodium cambricum (gauche): les trois niveaux de la succession coexistent sur quelques décimètres carrés. La fougère s'installe là où les deux premiers ont déjà préparé le terrain.
 Hiver
3 / 4
Hiver
3 / 4
Asplenium trichomanes (frondes à rachis brun, centre), Sedum sediforme glauque (bas), coussins de mousses autour. Le rhizome de la Doradille est ancré exactement là où Sedum et mousses ont accumulé quelques millimètres d' humus dans la fissure.
 Printemps
4 / 4
Printemps
4 / 4
Rosette en repos végétatif sur calcaire nu. À gauche, fronde desséchée de Polypodium cambricum avec sores apparents, les deux espèces partagent la même fissure de rocher. Mai 2026.
Sedum sediforme, l'Orpin de Nice, s'implante souvent en même temps que les premières fougères, une fois qu'une couche organique minimale s'est accumulée dans la fissure. Ses racines charnues ancrent l'humus naissant et l'empêchent de partir à la première pluie violente. En plein été, la partie aérienne ralentit sans disparaître: le stockage hydrique dans les feuilles succulentes compense l'aridité. Le réseau racinaire reste en place, et sa décomposition partielle ajoute chaque hiver une fraction organique supplémentaire dans la microfissure. Ce n'est pas une colonisation spectaculaire: l'Orpin de Nice consolide ce que les mousses ont mis des décennies à construire. Deux autres orpins ont été documentés sur le massif; leurs critères de détermination sont détaillés dans l'encart ci-dessus.
Ce que les pionniers préparent pour les fougères
Lichens
Dissolution chimique de la roche. Création d'une microrugosité de surface. Fixation d'azote atmosphérique.
Mousses
Rétention d'eau prolongée après la pluie. Accumulation de matière organique. Phosphore libéré sous formes labiles.
Sédums
Ancrage de l'humus contre l'érosion. Élargissement des fissures par les racines. Apport organique par décomposition hivernale.
Biofilm · stade précoce
Cyanobactéries et algues vertes sur roche nue. Microrugosité, fixation d'azote atmosphérique en faibles quantités, première rétention d'eau et de poussières. Invisible à l'œil nu, détectable aux traînées grises sur calcaire. Condition nécessaire à l'ancrage des premiers lichens crustacés.
Chlorolichens · stade précoce à tardif
Photobionte algue verte, se réactivent sur la seule vapeur d'eau atmosphérique. Les crustacés ( Verrucaria , Aspicilia ) ouvrent la voie dès le stade précoce; l'acide oxalique attaque CaCO₃ plus efficacement que le biofilm et libère les premiers minéraux dans les fissures. Les foliacés ( Xanthoria parietina ) suivent plus tard, là où une surface de rétention minimale s'est constituée: thalle décollable en partie, plus exposé mais plus efficace pour piéger les particules aéroportées.
Cyanolichens · stade médian à tardif
Collema , Lathagrium , Scytinium . Photobionte cyanobactérien ( Nostoc ), thalle gélatineux à l'état hydraté. Plus tardifs que les chlorolichens parce que leur photobionte exige de l'eau liquide pour réactiver la photosynthèse, indisponible sur roche nue. Fixent l'azote atmosphérique, contribution réelle au stock azoté du proto-sol.
Mousses · stade médian à tardif
Ptychostomum capillare , Tortella tortuosa . Coussins dans les fissures, rétention hydrique plusieurs jours, phosphore organique plus accessible que les apatites de la roche brute. Un sol de quelques millimètres se constitue à mesure que les coussins meurent et se renouvellent.
Fougères rupicoles · Doradille (médian-tardif) · Cétérach (tardif) · Polypode (tardif et au-delà)
La Doradille apparaît en premier, dans les fissures nord à sol fissural minimal: la géométrie de la fissure compense l'absence de substrat, par capillarité et condensation nocturne. Le Cétérach colonise les surfaces plus exposées et arrive ensuite. Sa tolérance à la dessiccation résout le problème de l'eau, pas celui du substrat: son rhizome reste plus volumineux que celui de la Doradille et exige quelques millimètres d'accumulation interstitielle pour s'ancrer. Le Polypode, le plus exigeant en matière organique, suit en dernier: il a besoin de l'humus que les mousses ont mis des décennies à constituer. Les trois peuvent coexister sur la même paroi à des stades différents, décalés de quelques décimètres.
Durées extrapolées d'après Belnap & Lange, 2001 et données de colonisation sur murs calcaires Darlington, 1981 . Aucune chronométrie directe n'a été effectuée sur le massif de la Clape.
Lichens, mousses et sédums occupent la roche longtemps avant l'arrivée d'une fougère. Quand un sporophyte de Polypode ou de Cétérach s'installe vers T+30 à T+40, il bénéficie d'une infrastructure déjà constituée: substrat micro-rugueux entretenu par les lichens, réserve d'eau dans les coussins de mousse, proto-sol enrichi par les sédums. Chacun des trois voisins apporte quelque chose de différent et complémentaire. Aucun n'attend de retour: la facilitation est strictement unidirectionnelle, le plus simple ouvre le passage au plus complexe.
Le lichen: préparation du substrat
Échelle: décennies, T+5 à T+30 environ
Les lichens, surtout les chlorolichens crustacés ( Aspicilia , Verrucaria ), dissolvent microscopiquement le carbonate de calcium par sécrétion d'acide oxalique. À mesure que le thalle vit puis meurt, il creuse une microrugosité de 50 à 200 µm et dépose une nécromasse organique qui forme la première couche de proto-sol exploitable. Les particules minérales aéroportées (sables, poussières) s'y piègent.
Pour les cyanolichens spécifiquement ( Collema , Lathagrium , Scytinium ), ajouter un apport d'azote fixé via leur photobionte cyanobactérien. Les chlorolichens, eux, ne fixent pas d'azote : la distinction est importante, tous les lichens ne sont pas équivalents sur ce plan.
La mousse: l'eau pour la reproduction
Échelle: semaines à mois, à un moment précis du cycle fougère
Les coussins de mousses retiennent typiquement trois à dix fois leur masse sèche en eau, parfois plus chez les formes pulvinées denses, et la libèrent lentement par évaporation depuis les capillaires interfeuillets Proctor, 2008 Glime, 2017, ch. 7 . Les mousses xérophytes calcicoles présentes sur la Clape ( Tortella tortuosa , Schistidium apocarpum , Ptychostomum capillare , Grimmia pulvinata ) sont plutôt dans la partie basse de cette fourchette, les sphaignes des tourbières qui dépassent vingt fois leur masse sèche en eau étant absentes du massif. Apports secondaires : matière organique, ombrage, tamponnage thermique, phosphore organique plus accessible que les apatites de la roche brute.
L'apport critique se joue sur la phase invisible du cycle fougère. Quand une spore germe sur ou dans un coussin de mousse, elle donne un prothalle , le gamétophyte de la fougère: un petit organisme de quelques millimètres, sans cuticule ni racine, qui doit survivre plusieurs mois en milieu humide. La fécondation exige un film d'eau pour la nage des anthérozoïdes multiflagellés vers l'archégone ; le jeune sporophyte exige une humidité maintenue avant que ses racines ne soient fonctionnelles. Sans mousse, pas de prothalle viable. Sans prothalle, pas de génération suivante. C'est la dépendance la plus forte de tout le système, et elle est invisible: on regarde une touffe adulte qui semble vivre sa vie, sa génération suivante dépend pourtant de ce que fait la mousse à quelques centimètres.
Le sédum: proto-sol sans compétition pour l'eau
Échelle: décennies, T+10 à T+50+
Les sédums présents sur la Clape ( Sedum sediforme , S. acre , Sedum gr. rupestre s.l.) ont des feuilles charnues et un renouvellement de biomasse abondant. En mourant, ils s'accumulent dans les anfractuosités et se décomposent en une mince couche organo-minérale: c'est le proto-sol , quelques millimètres à quelques centimètres de matière sans horizons différenciés, juste assez pour qu'une spore puisse germer. Leur décomposition produit la fraction la plus organique du proto-sol des fissures, plus substantielle en masse que celle des lichens ou des mousses, et restitue K, Mg, Ca au système.
Particularité métabolique décisive: Sedum sediforme et S. acre pratiquent la photosynthèse CAM ( Crassulacean Acid Metabolism ), inverse du fonctionnement des fougères ; le statut de Sedum gr. rupestre s.l. dans ce complexe est moins établi. Leurs stomates s'ouvrent la nuit, quand l'air est plus humide et la température basse ; ils restent fermés le jour. Conséquence: pour fixer une même quantité de carbone, un sédum CAM perd deux à cinq fois moins d'eau qu'une plante en photosynthèse C3 standard. Il n'épuise pas l'humidité disponible à la fougère voisine. Les niches sont par ailleurs partiellement disjointes: sédums sur micro-corniches ensoleillées bien drainées, fougères dans fissures ombragées plus profondes. Cohabitation plus que compétition.
Une vraie symbiose existe pour la fougère, mais ailleurs. Le voisinage de surface est unidirectionnel: lichens, mousses et sédums donnent sans recevoir. La symbiose mutualiste de la fougère, elle, se joue sous la roche , dans le rhizome. Les fougères calcicoles entretiennent presque toutes des associations mycorhiziennes arbusculaires: des champignons Glomeromycota colonisent les cellules du cortex racinaire et étendent par leurs hyphes une zone d'absorption d'eau et de phosphore mille à dix mille fois plus large que celle des racines seules. C'est cette association qui constitue la symbiose centrale d'une fougère rupestre, invisible parce qu'enfouie. Voir le développement détaillé au chapitre suivant, dans la section consacrée aux associations mycorhiziennes .
Ce système n'est donc pas un réseau mutualiste, c'est un système de facilitation unidirectionnelle où les organismes les plus simples préparent les conditions des plus complexes sans attente de retour. L'absence de réciprocité n'est pas une lacune: c'est le mode de fonctionnement normal d'une succession écologique en milieu contraignant. La fougère arrive à la fin de la chaîne, elle hérite, elle ne rend rien aux étages précédents qu'une ombre légère et de la litière modeste, et c'est suffisant.
Ce mécanisme porte un nom en écologie végétale.
La théorie du gradient de stress ( SGH )
Stress Gradient Hypothesis · Bertness & Callaway, 1994
En milieu favorable, les plantes se font surtout concurrence pour la lumière et les nutriments. En milieu hostile, les interactions positives entre espèces, la facilitation , deviennent dominantes. Plus le milieu est stressant, plus les espèces s'entraident plutôt qu'elles ne se concurrencent. Dans la Clape, lichens, mousses, sédums et fougères forment une guilde pionnière où chaque groupe crée les conditions qui permettent au suivant de s’installer. Cette hiérarchisation vaut pour les interactions inter-strates (lichens → mousses → fougères). Les interactions intra-strates, notamment entre espèces de lichens sur une même surface, restent moins documentées et peuvent inclure une part de compétition même en milieu très contraint Maestre et al., 2009 .
La facilitation pionnière prépare le terrain, les refuges microclimatiques créent les conditions locales. Ces contraintes générales posées, il reste à comprendre ce que les trois fougères font à leur tour, en commun puis chacune à sa manière, pour s'y maintenir: c'est l'objet de la partie suivante.
Les mécanismes
Les trois espèces partagent un même point de départ: un substrat sans sol, une eau intermittente, et au niveau du rhizome à peu près les mêmes mécanismes pour mobiliser les nutriments bloqués par le calcaire. Au-dessus du sol, les réponses à la sécheresse divergent franchement. Cette partie commence par ce qui leur est commun, puis examine ce qui distingue chacune: la tolérance physiologique du Cétérach, la dormance saisonnière du Polypode, le choix d'habitat de la Doradille. Ces réponses ne s'excluent pas. Chaque espèce recourt aussi, à un moindre degré, à ce que les deux autres font en priorité.
✦ ✦ ✦
Avant que chaque espèce ne déploie sa stratégie propre face à la sécheresse, les trois font face au même problème de fond : sur un substrat calcaire à pH élevé, le phosphore est présent dans la roche mais chimiquement bloqué, quasi inaccessible pour les racines. C'est le verrou nutritif partagé, celui que chacune doit lever avant tout le reste. Le phosphore intervient à quatre niveaux cellulaires fondamentaux : il constitue le squelette de l'
ADN
et de l'
ARN
, assure le transfert d'énergie via l'
ATP
, et entre dans la composition des
phospholipides
constitutifs de toutes les membranes cellulaires.
L'ARN à lui seul représente la moitié ou plus du phosphore non stocké d'un organisme photosynthétique
Raven, 2013
, si bien que la demande se concentre dans les tissus à division rapide:
méristèmes
du rhizome, jeunes frondes et structures reproductrices, où la synthèse des acides nucléiques est la plus intense. Une carence en phosphore freine donc d'abord la croissance et le renouvellement, fonctions sur lesquelles repose tout le cycle. Le phosphore est le mieux documenté de ces verrous, et le plus aigu: c'est sur lui que portent les mécanismes décrits ci-dessous. C'est un préalable physiologique partagé, sur lequel les divergences plus visibles vont ensuite s'appuyer.
Une clé de lecture : la théorie CSR
Compétiteurs · Tolérants au stress · Rudéraux · Grime, 1977
Les plantes se répartissent en trois grandes stratégies selon leur rapport au stress et aux perturbations. Les compétiteurs (C) poussent vite et accaparent les ressources en milieu favorable. Les rudéraux (R) colonisent rapidement les milieux perturbés. Les tolérants au stress (S) ont une croissance lente, une longue durée de vie et des besoins réduits: ils persistent là où les autres s'épuisent. Sur une paroi calcaire méditerranéenne, à stress élevé et perturbations faibles, ce sont les stratèges S qui restent. Les trois fougères de la Clape correspondent à ce profil: croissance lente, faible emprise au sol, présence maintenue là où les plantes à croissance rapide disparaissent à la première sécheresse. Ce qui suit détaille les mécanismes partagés de ce profil, puis les variantes propres à chaque espèce.
Le verrou chimique du calcaire bloque deux éléments en priorité : le phosphore, précipité avec le calcium en apatite insoluble, et le fer, dont la solubilité du Fe³⁺ s'effondre dès que le pH dépasse 7 (environ six ordres de grandeur entre pH 6 et pH 8 Lindsay, 1979 ). Le mécanisme détaillé et les fractions concernées sont décrits dans La chimie du calcaire au chapitre précédent. Les deux autres macronutriments majeurs ne sont pas dans la même situation.
Pourquoi pas l'azote ? Sur calcaire, l'azote n'est pas le verrou principal. La nitrification bactérienne fonctionne à pH alcalin, et les cyanobactéries fixatrices du biofilm épilithique apportent l'azote atmosphérique sur la roche. La matière organique accumulée dans les fissures en libère aussi par minéralisation. Les fougères rupestres de la Clape ne présentent pas la symptomatologie de carence azotée, qui produirait un jaunissement diffus et uniforme de l'ensemble du feuillage, sans lien avec le pH.
Pourquoi pas le potassium ? Le potassium est libéré par l'altération des silicates et reste mobile en solution à pH alcalin : il ne précipite pas avec le calcium dans les conditions du massif. Les carences potassiques existent sur certains calcaires, mais elles ne constituent pas le facteur limitant pour des rhizomes courts qui extraient leurs nutriments de volumes de fissure restreints sur un substrat minéralement riche.
Autour du rhizome, sur quelques millimètres, la fougère modifie localement la chimie pour décrocher les nutriments que le calcaire retient. Pour le phosphore, elle combine une acidification du sol et des acides organiques qui captent le calcium et libèrent le phosphate piégé. Pour le fer, elle le rend soluble en le réduisant à la surface de ses racines, une voie qui reste efficace en milieu alcalin mais s'épuise quand le pH monte trop : c'est pourquoi le jaunissement des feuilles par manque de fer, la chlorose ferrique , reste fréquent sur calcaire. Le détail biochimique des deux voies figure ci-dessous.
Sur le ciste cotonneux et les cistes du massif: Exploration & histoire de Cistes · Brûlures et renaissances , maclape.com. Sur la biologie et l’écologie du genévrier de Phénicie dans le massif: Flora Verticalis Part. I · Genévriers de la Clape , maclape.com. Sur le caprifiguier enraciné dans la même roche: Figuiers sauvages de la Clape , maclape.com.
Les trois espèces forment vraisemblablement des associations mycorhiziennes à arbuscules ( AMF ), documentées chez les fougères leptosporangiées en général et partiellement confirmées pour les genres Asplenium et Polypodium sur calcaire Turnau et al., 2005. Ces champignons étendent la prospection au-delà des micropores inaccessibles aux radicelles et sécrètent des phosphatases qui libèrent le phosphore organique accumulé dans les fissures. Dans un substrat aussi pauvre en phosphore disponible que les lapiaz de la Clape, cette association pèse sur l'établissement de jeunes sporophytes, dont la survie dépend d'un accès précoce aux nutriments. Les données disponibles sur les populations saxicoles méditerranéennes restent fragmentaires: l'encart ci-dessous précise ce qui est établi et ce qui reste à documenter.
Ce travail souterrain, les trois espèces le mènent à peu près de la même façon. Au-dessus, en revanche, les divergences sont nettes: ce qui distingue le Cétérach, le Polypode et la Doradille, c'est leur manière respective de tenir face à la sécheresse.
En plein été, les frondes du Cétérach sont roulées sur elles-mêmes, brunies, friables. On le croirait mort. Ce n'est pas le cas. C'est l' anhydrobiose , forme extrême de tolérance à la dessiccation: la plante se déshydrate jusqu'à atteindre une teneur en eau relative ( RWC ) résiduelle de 4 à 7 %, soit 93 à 96 % d'eau perdue Breuil, 1997 ; Proctor, 2009 . Son métabolisme s'interrompt, elle attend. Dès les premières pluies d'automne, en 2 à 4 h en conditions expérimentales Proctor, 2009 , elle se réhydrate et reprend une couleur verte normale. Le profil des composés volatils émis au cours de cette reprise est documenté dans Živković et al., 2021 .
Cette capacité à tolérer la dessiccation quasi-totale est peu répandue chez les plantes vasculaires. Elle repose sur des mécanismes moléculaires précis, les mêmes que ceux qui protègent les graines des plantes à fleurs : le Cétérach les applique non pas à une graine mais à l'ensemble de ses cellules végétatives actives, ce qui est beaucoup plus rare.
Ces mécanismes ne fonctionnent pas seuls. La morphologie de la fronde s'y ajoute: la face inférieure couverte d'écailles apporte une protection physique qui double la résistance biochimique interne.

Les paléas , décrites dans le portrait de Observer pour comprendre , ne sont pas un simple ornement. En été, quand les frondes se roulent sur elles-mêmes, elles se retrouvent exposées vers l'extérieur: elles réfléchissent une partie du rayonnement solaire et réduisent l'échauffement de la fronde. En période humide, elles forment une barrière physique qui réduit la transpiration cuticulaire et ralentit l'évaporation. Ce double rôle, protection thermique et régulation hydrique, s'active successivement selon la saison.
 Hiver
1 / 6
Hiver
1 / 6
Paléas imbriquées sur la face inférieure de la fronde. Les écailles semi-transparentes laissent affleurer les sporanges sombres en dessous.
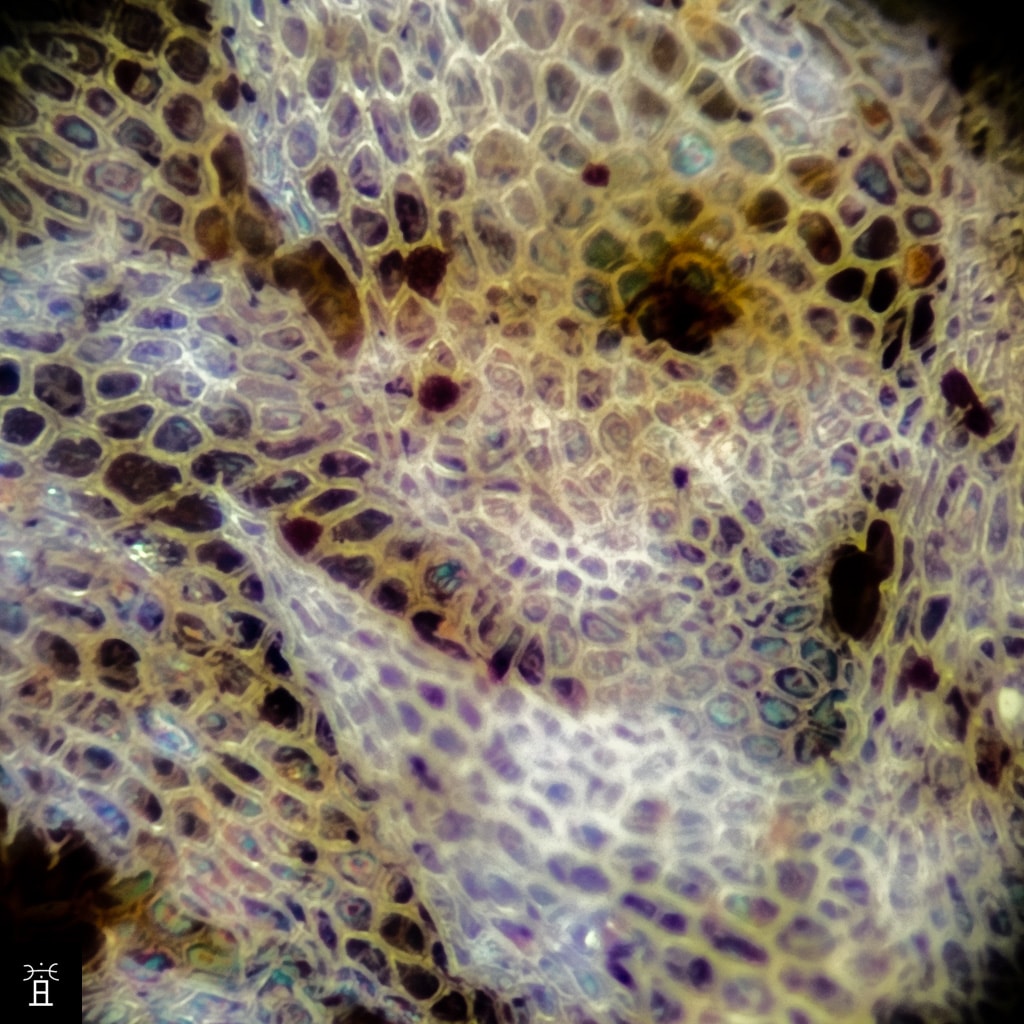 Hiver
2 / 6
Hiver
2 / 6
Structure clathrée des écailles de la face inférieure: les parois cellulaires épaissies forment un réseau alvéolaire dense. Ce feutrage imperméabilise la fronde et réfléchit une partie du rayonnement solaire quand les frondes se roulent en été.
 Printemps
3 / 6
Printemps
3 / 6
Sur les parois les plus exposés, ce Cétérach va bientôt entrer dans sa phase de protection et de dormance estivale. Les crosses encore étroitement enroulées et protégées par leurs écailles protectrices
 Été
4 / 6
Été
4 / 6
Logé dans une fissure horizontale, dès que la ressource en eau diminue, les frondes du Cétérach se roulent sur elles-mêmes jusqu'à paraitre mortes (Juin 2026).
 Été
5 / 6
Été
5 / 6
Frondes entièrement enroulées, recouvertes de paléas brun-doré imbriqués. La structure en tire-bouchon est caractéristique d'une dessiccation poussée : chaque fronde s'est vrillée sur elle-même pour réduire la surface exposée au rayonnement.
 Été
6 / 6
Été
6 / 6
Détails des Frondes: On distingue encore, dans les replis internes, des zones vert sombre : le tissu chlorophyllien est intact et dormant, non nécrosé. La reprise sera complète dès les premières pluies.
Ce double dispositif, biochimique et morphologique, ouvre au Cétérach une niche que les deux autres espèces de la Clape ne peuvent pas occuper.
Le Cétérach pousse là où les deux autres espèces ne vont pas: sur les parois exposées, en plein soleil. Sa tolérance à la dessiccation lui ouvre ces stations. La seule contrainte réelle: quelques millimètres de sol dans la fissure pour ancrer son rhizome.
Le Cétérach tient parce qu'il résiste. L'espèce suivante a trouvé une réponse entièrement différente au même problème: ne pas être là quand la sécheresse arrive. Là où le Cétérach résiste, le Polypode esquive.
 Printemps
Printemps
Déhiscence printanière · Mars
 Printemps
1 / 4
Printemps
1 / 4
Fronde en déploiement: pennes inférieures étalées avec sores orange mûrs, pennes supérieures encore en crosse. Les deux stades coexistent sur la même fronde. Déhiscence imminente.
 Printemps
2 / 4
Printemps
2 / 4
Sores orange-brun matures, déhiscence en cours. Les frondes s'enroulent: le rhizome amorce son retrait. Sporulation et entrée en dormance coexistent sur le même pied.
 Été
3 / 4
Été
3 / 4
Frondes desséchées, tordues, brun-jaune. Sores vides encore visibles en points sombres. La plante paraît morte: le rhizome est intact. Elle reprendra en octobre.
 Été
4 / 4
Été
4 / 4
Frondes complètement desséchées. Elles vont bientôt se détacher du rizhome.
Polypodium cambricum adopte une stratégie radicalement différente: il ne résiste pas à la sécheresse, il l'évite par le temps. À partir de juin, le rhizome réduit ses échanges gazeux, concentre ses réserves glucidiques et suspend l'élongation cellulaire. Les frondes se dessèchent et tombent, le rhizome reste intact. Il ne reste rien de visible: la hausse des températures et la baisse de l’humidité ont induit une dormance physiologique. Dès les premières pluies d'automne, la croissance reprend. Les nouvelles frondes apparaissent en quelques semaines, la plante est en pleine activité photosynthétique en hiver, ses spores sont mûres au printemps. Cette plasticité phénologique est sa force principale. Ce type d' estivation est commun chez les bulbes et les géophytes méditerranéens ; il est rare chez les fougères des flores tempérées d’Europe continentale, et mieux documenté dans les zones méditerranéennes et subtropicales où d’autres Polypodiacées ont développé des stratégies comparables.
Ce calendrier inversé n'est pas seulement une esquive de la sécheresse. Il exploite aussi une fenêtre de ressources que les autres plantes laissent libre.
En hiver, dans les stations à exposition nord où se développe le Polypode, les journées sont courtes et la lumière reste diffuse, ce qui limite le risque de photo-inhibition . L'eau est disponible. Les températures restent la plupart du temps au-dessus de zéro dans les stations du massif, et les gels brefs et peu intenses qui peuvent survenir sont tolérés par les frondes actives. Sur la longue série Narbonne, le nombre annuel de jours de gel a reculé de 22 jours dans les années 1960-1970 à 9 jours sur la période 2000-2024, soit une division par deux et demie. La contrainte gélive, jamais centrale pour cette espèce, devient encore plus marginale. Le Polypode profite d'une fenêtre temporelle que les plantes annuelles d'été laissent libre. C'est une forme d'évitement par le temps, au lieu de l'évitement dans l'espace que pratique la Doradille.
 Printemps
Printemps
Rhizome et accumulation organique · Mai
En hiver, ce qu'on voit sur la paroi c'est du vert. Ce qu'on ne voit pas, c'est le rhizome rampant couvert d'écailles brunes, serré contre la roche, qui a produit un lacis de racines adventives ancrées dans les diaclases du calcaire. Ce réseau ne disparaît pas avec les frondes. Pendant que celles-ci sèchent et tombent en juin, le rhizome reste en place. C'est lui qui porte les réserves glucidiques qui rendront possible la reprise d'octobre.
Sur les pieds bien établis, ce tapis d'écailles et de racines retient les poussières éoliennes et les débris organiques déposés par les ruissellements. Une couche de matière organique fine s'installe entre la plante et la roche, quelques millimètres au plus. Elle ralentit le séchage du substrat après chaque pluie et constitue une réserve de nutriments là où la roche nue n'en offre aucune. En années sèches, quand les précipitations du massif tombent entre 300 et 400 mm, ce différentiel de rétention est mesurable. Son rôle précis pour la survie du rhizome dans les populations saxicoles de la Clape reste à quantifier.
 Printemps
Printemps
Réseau racinaire adventif Polypodium cambricum · Mars
Le Polypode esquive la sécheresse dans le temps. La troisième espèce résout le même problème dans l'espace. Elle ne dort pas, elle ne résiste pas: elle choisit simplement l'endroit où la sécheresse ne l'atteint pas.

Asplenium trichomanes n'a ni la résistance biochimique du Cétérach ni la souplesse phénologique du Polypode. La Doradille ne résiste pas à la sécheresse et ne l’esquive pas non plus. Elle s’installe là où elle n’a pas à le faire: dans les fissures profondes où la roche garde un peu d’humidité même en juillet. C’est une stratégie spatiale, pas physiologique.
Une clé de lecture : la théorie des refuges microclimatiques
Microclimate refuge theory · Ashcroft, 2010
Certaines zones de petite taille présentent des conditions thermiques et hydriques qui s'écartent significativement des conditions régionales. Ces refuges permettent à des espèces d'y persister alors que le milieu environnant leur est devenu hostile. Dans la Clape, les fissures orientées au nord fonctionnent ainsi pour la Doradille capillaire: la température y reste de quelques degrés inférieure à celle des parois exposées, l'humidité plus stable, et cet écart de quelques centimètres suffit à changer ce qui est biologiquement possible. C'est exactement la stratégie spatiale décrite ci-dessus.
Les conditions qu'elle exploite tiennent aux deux mécanismes décrits au cycle de l'eau dans Le cadre environnemental : la capillarité dans les interstices étroits, qui retient un film d'eau plusieurs jours après la pluie, et la condensation nocturne sur les parois nord, qui en renouvelle quelques dixièmes de millimètre chaque matin. Ce microclimat de quelques centimètres carrés suffit à changer ce qui est biologiquement possible pour une fougère qui ne peut ni résister ni esquiver la sécheresse.
 Hiver
1 / 3
Hiver
1 / 3
Vue plongeante dans une concavité orientée nord. La Doradille occupe le fond, encadrée de mousses. La géométrie dit tout: la Doradille choisit le creux plutôt que la paroi, parce que les conditions y sont différentes.
 été
2 / 3
été
2 / 3
Touffe d'Asplenium trichomanes dans une fissure de calcaire, juin 2026. Rachis brun-noir arqués et retombants, pinnules réduites et recourbées vers le bas : réponse thermonastique à la chaleur estivale. La plante réduit sa surface évapotranspirante sans entrer en dormance, contrairement au Cétérach, elle ne tolère pas la dessiccation complète et maintient un minimum de turgescence.
 été
3 / 3
été
3 / 3
Détail des rachis et pinnules en juin: chaque pinnule s'enroule sur elle-même, face adaxiale concave, bords relevés en gouttière. Ce mouvement réduit l'exposition directe au rayonnement solaire et limite les pertes en eau par transpiration.
Choisir la fissure n'est pas simplement un comportement passif de dispersion: la forme de la plante elle-même est adaptée à y vivre. La Doradille ne se contente pas de tomber dans une fissure par hasard de spore, sa morphologie la rend particulièrement efficace dans ce milieu.
Le rachis brun foncé décrit dans Observer pour comprendre n'est pas seulement un critère de détermination: sa forte lignification lui confère une résistance à la dessiccation que les pinnules n'ont pas. Quand celles-ci tombent, lui reste en place, signalant la présence d'un individu même en pleine sécheresse.
La forme même de la fronde est adaptée à la vie dans les interstices: longue et fine, elle progresse dans la fissure sans avoir besoin d'espace large. Certains individus poussent presque horizontalement, leurs frondes s'avançant dans l'interstice comme si elles cherchaient quelque chose.
L'eau dans une fissure étroite est tirée vers le haut par les forces d'adhésion entre le liquide et la roche: plus la fissure est fine, plus ces forces dominent le poids de la colonne d'eau et plus la remontée est haute. Une fissure de 5 mm retient un film d'eau sur quelques millimètres. Une fissure de 0,5 mm le retient sur plusieurs centimètres, parfois davantage. La Doradille s'installe dans la seconde catégorie, là où la physique fait un travail que la plante n'a pas à faire elle-même. Sa morphologie répond exactement à cette dimension: pinnule de quelques millimètres, rachis filiforme, fronde rarement plus large qu'un crayon. Une plante plus charnue ne pourrait pas s'y loger ; une plante plus haute n'y trouverait plus l'humidité qu'elle exploite.
Cette géométrie a un autre avantage. Quand le film capillaire se rétracte en été, le rhizome reste en contact avec la roche encore humide en profondeur, plus loin dans la fissure. Les radicelles latérales de la Doradille sont plus développées que la partie aérienne ne le laisse deviner ; elles colonisent l'interstice horizontalement, là où une racine pivotante ne pourrait pas suivre. La plante visible en surface n'est qu'une fraction de l'individu installé dans la roche.
Les trois espèces ont été décrites séparément, chacune par sa réponse dominante. Les stratégies se recoupent pourtant. Le Cétérach tolère la dessiccation, mais il sélectionne aussi son habitat et ralentit son métabolisme en été. Le Polypode esquive par la phénologie, mais il occupe des expositions précises et tolère de brefs gels hivernaux. La Doradille mise sur la fissure, mais son rachis lignifié et ses pinnules légèrement hydrophobes lui confèrent une résistance partielle à la dessiccation. Les mécanismes sont les mêmes pour les trois ; c'est leur hiérarchie qui diffère. Le tableau ci-dessous résume les paramètres qui les distinguent.
Les trois espèces côte à côte
Comparaison des paramètres écologiques et adaptatifs
| Paramètre | Cétérach | Polypode austral | Doradille capillaire |
|---|---|---|---|
| Stratégie | Tolérer la dessiccation | Esquiver par le temps | Se glisser dans l'espace |
| Saison active |
Automne–printemps En veille l'été |
Automne–hiver–printemps Dormant l'été |
Toute l'année Ralentie l'été |
| Habitat préférentiel | Parois et lapiaz, toutes expositions | Falaises et blocs orientés nord | Fissures étroites et profondes |
| Mécanisme clé | Anhydrobiose + paléas argentés | Dormance estivale du rhizome | Microclimat de fissure |
Comment les trois stratégies se combinent dans chaque espèce
|
Tolérance anhydrobiose |
Esquive temporelle dormance |
Refuge spatial fissure |
|
|---|---|---|---|
| Cétérach | dominante | ralentissement métabolique estival | ancrage fissural minimal |
| Polypode | tolérance partielle au déficit hydrique | dominante | exposition nord, écailles du rhizome |
| Doradille | rachis lignifié, pinnules hydrophobes | ralentissement estival par perte des pinnules | dominante |
dominante structurant la biologie de l'espèce · secondaire, marge d'ajustement. Les trois mécanismes coexistent dans chaque espèce ; ce qui les distingue est leur hiérarchie.
Ces trois stratégies se complètent. Dans le massif, Cétérach et Doradille se retrouvent régulièrement dans la même fissure, parfois à quelques centimètres l'un de l'autre. La tentation est de parler de compétition . Ce n'en est pas. Ce que l'on voit, c'est deux espèces qui occupent des portions différentes d'un même espace, sans que leurs exigences se recoupent.
 Mai
Mai
Cétérach et Doradille dans la même fissure
Le Cétérach tient le bord exposé de la fissure: la lèvre supérieure, parfois la paroi verticale entre deux interstices. C'est là que le rayonnement solaire arrive directement en été. La Doradille, elle, s'enfonce dans la fissure elle-même. Ses frondes émergent de l'interstice, mais son rhizome est ancré là où la roche maintient un film d'humidité que la surface ne conserve pas.
Cette limite entre les deux espèces correspond souvent à un écart thermique de 5 à 8 °C en pleine journée de juillet/août. Le Cétérach sur les bords exposés peut dépasser 50 °C en surface pendant les heures les plus chaudes. Il supporte cela parce qu'il est déjà passé en anhydrobiose, état déshydraté stable installé progressivement au printemps avant les pics thermiques. La Doradille, à 4 cm de profondeur, ne dépasse pas 38 °C et conserve une humidité capillaire résiduelle. Elle ne tolère pas la dessiccation: elle n'a pas à la tolérer.
La cohabitation dans la même fissure pose une question directe: les deux espèces peuvent-elles s'hybrider ? La réponse est non. Aucun hybride entre A. ceterach et A. trichomanes n'a jamais été décrit dans la littérature, et la phylogénie explique pourquoi : les deux espèces appartiennent à des clades majeurs distincts au sein du genre Asplenium , le « Phyllitis clade » (II) pour le Cétérach et le « A. trichomanes clade » (VI) pour la Doradille Xu et al., 2020 . Les deux espèces présentes dans la Clape sont tétraploïdes (2n = 144 chez le Cétérach, 2n = 144 chez la Doradille subsp. quadrivalens ), mais cette identité numérique de ploïdie ne suffit pas : ce sont des génomes issus d'ancêtres différents. Dans la Clape, leurs spores germent parfois sur le même substrat, leurs prothalles coexistent, l'eau de condensation les atteint au même moment. Rien de cela n'a produit d'hybride, ici ni ailleurs en Méditerranée. La séparation entre ces deux clades remonte au rayonnement initial du genre Asplenium au Paléocène, daté entre 55 et 60 Ma par horloge moléculaire Schneider et al., 2004 ; Schuettpelz & Pryer, 2009 ; aucune cohabitation, même prolongée par la mise en place du climat méditerranéen au Miocène-Pliocène, n'a suffi à la franchir.
Toutes trois partagent pourtant une contrainte que ces partitions spatiales et temporelles ne font pas disparaître: la fécondation exige de l'eau, au même moment, pour les trois. C'est l'objet de la partie suivante.
Synthèse
Trois façons de tenir dans un endroit qui ne vous a pas attendu. Mais tenir en tant qu'individu ne suffit pas: le cycle exige de l'eau à un moment précis, et aucune de ces adaptations ne peut contourner cette contrainte.
Le cycle
Le Cétérach reprend en quelques heures après la pluie. Le Polypode repart en octobre. La Doradille ne s'arrête presque pas. Mais aucune des trois n'a résolu le problème de la fécondation autrement que ses ancêtres du Carbonifère: il faut de l'eau, un film sur le prothalle, au bon moment.
✦ ✦ ✦
La fronde est un organe saisonnier ou bisaisonnier ; le rhizome est l'organe de continuité. Les trois espèces convergent là-dessus : chez le Polypode, il reste intact pendant toute la dormance estivale ; chez le Cétérach, une pluie d'automne de 20 mm au moins suffit à relancer l'activité depuis la veille du rhizome Proctor, 2009 ; chez la Doradille, les rachis lignifiés persistent bien après la chute des pinnules et signalent sa présence. Sa durée d'occupation se mesure différemment selon l'espèce : longueur cumulée pour le Polypode dont le rhizome rampe, densification des frondes superposées pour le Cétérach dont le rhizome reste court, accumulation des rachis morts pour la Doradille. Ces trois registres conduisent à des estimations chiffrées, plus solides pour le Polypode que pour les deux Asplenium .
Chez les plantes à fleurs, le cycle passe par une fleur, un pollen, une graine. Chez les fougères, c'est plus long. Il y a deux phases successives, deux plantes pour ainsi dire, l'une issue de l'autre.
La plante que l'on voit et reconnaît sur le terrain, avec ses frondes et son rhizome, est le sporophyte . C'est la génération principale, diploïde (deux jeux de chromosomes, noté 2n). Cette diploïdie n'est pas un détail génétique: elle est le prérequis du cycle. Pour produire des spores, les cellules mères au sein des sporanges du sporophyte doivent passer par la méiose , une division cellulaire réductrice qui ramène de 2n à n. Sans diploïdie en point de départ, pas de méiose possible, donc pas de spores haploïdes. Ces spores, microscopiques, sont ce que le vent emporte. La fécondation, plus loin dans le cycle, restaurera le niveau diploïde en fusionnant deux cellules haploïdes: n + n = 2n. C'est cette alternance entre les deux états qui constitue le cycle, et non simplement les formes visibles qu'il produit.
Les spores sont disséminées par le vent (anémochorie). Elles peuvent parcourir de longues distances. Leur paroi externe, constituée de sporopollénine , les protège contre la dessiccation, les rayons UV et les variations chimiques. Une spore peut rester viable plusieurs années. C'est précisément cette résistance qui leur permet d'atteindre des fissures inaccessibles à n'importe quelle graine.
Le mécanisme de libération est remarquable par sa précision : les spores sont éjectées par temps sec, exactement quand le vent peut les emporter le plus loin. C'est l'abaissement de l'humidité lui-même qui déclenche le tir.
 Hiver
Hiver
Vue macro des sporanges et annulus de l' Asplenium ceterach · Janvier

Sporanges du Polypode à différents stades · Janvier à Juin
La libération des spores se fait donc par temps sec, quand le vent peut les emporter. La germination et la fécondation, elles, exigent de l'humidité. Une fois la spore arrivée dans une fissure, elle ne donne pas immédiatement un prothalle: elle commence par gonfler par imbibition, puis s'ouvre et émet un filament de quelques cellules avant que la lamelle cordiforme se mette en place. Cette phase de germination proprement dite dure plusieurs semaines — durée précise non documentée ici pour les trois espèces en conditions méditerranéennes. C'est la période la plus exposée du cycle: plus petite que le prothalle lui-même, sans aucune protection, elle disparaît sans laisser de trace au moindre assèchement. C'est sur le prothalle constitué que se joue la suite.
Quand une spore germe, elle ne donne pas directement une nouvelle fougère. Elle produit d'abord une structure intermédiaire: le prothalle (ou gamétophyte ). C'est une lamelle verte de quelques millimètres, en forme de cœur aplati, qui adhère au substrat. Contrairement à ce qu'on pourrait croire, ce prothalle n'est pas un stade de la fougère: c'est un organisme à part entière, haploïde, qui vit, photosynthétise et meurt indépendamment. La fougère que vous voyez sur la paroi, le sporophyte, produit les spores par méiose — c'est sa façon de se reproduire, mais par voie asexuée. La reproduction sexuée appartient au prothalle: c'est lui qui porte les gamètes et assure la fécondation, pendant les quelques semaines à quelques mois où il survit. Dans la Clape, c'est aussi l'organisme le plus vulnérable de tout le cycle: 2 à 5 mm, pas d'écailles, pas de rhizome, aucune des protections que le sporophyte a développées. Un seul épisode de sécheresse au mauvais moment l'élimine avant la fécondation. C'est sur ce prothalle que se forment les organes sexuels, et c'est là qu'intervient la contrainte décisive.
Le point de passage obligé: la fécondation
Les gamètes mâles des fougères, les anthérozoïdes , sont mobiles et portent de nombreux flagelles (multiflagellés). Pour rejoindre l'organe femelle (l' archégone ) et féconder l'oosphère, ils doivent nager dans un film d'eau liquide à la surface du prothalle.
Pas d'eau, pas de fécondation. Un épisode de rosée matinale, une pluie légère après une nuit de condensation: c'est souvent tout ce qu'il faut, mais ce minimum est indispensable. Dans un massif méditerranéen, c'est le goulot d'étranglement du cycle.
Tous les autres végétaux vasculaires du massif (genévrier de Phénicie, ciste, figuier) s'affranchissent de cette contrainte via le pollen. Les fougères, non.
Le moment où cette fenêtre humide se présente n'est pas le même pour les trois espèces : il est détaillé dans Les fenêtres de reproduction dans la Clape .
Cycle de reproduction d'une fougère, les deux générations
Échelle de taille des structures, du micron au mètre
Le cycle ci-dessus montre la double contrainte parcourue : la dispersion des spores demande de la sécheresse et du vent, la fécondation exige un film d'eau liquide sur le prothalle. Ces deux exigences contradictoires, qui ne coexistent jamais au même moment dans la Clape, structurent le calendrier reproductif des fougères.
Dans le massif, les trois espèces ont décalé leur période de fécondation vers les moments où l'eau est disponible: l'automne surtout, et le début du printemps. Mais elles n'ont pas le même calendrier exact.
 Printemps
Printemps
Cétérach, chevauchement des cycles sur une même touffe · Mai
Cétérach
Asplenium ceterach
Spores matures en fin d'été. Libération en août-septembre par temps sec: l'annulus se rétracte par évaporation et éjecte les spores, le mécanisme est actif pendant la sécheresse, non à son début. Germination à l'automne sur parois réhydratées par les premières pluies. Fécondation: automne à début hiver.
Polypode austral
Polypodium cambricum
Sores visibles de novembre à mars sur frondes actives. Spores libérées en hiver-printemps. Germination au printemps suivant avec l'humidité printanière. Cycle le plus cohérent avec les pluies hivernales méditerranéennes.
Doradille capillaire
Asplenium trichomanes
Double période de sporulation et de libération des spores: avril–mai et septembre–octobre. Le microclimat de la fissure maintient une humidité suffisante pour la germination même en période relativement sèche. Fécondation possible dès les premières condensations nocturnes.
Calendrier de reproduction
Les trois espèces · Massif de la Clape
Synthèse
Le cycle des fougères pose une équation en deux termes: la dispersion des spores veut du vent sec, la fécondation veut de l'eau. Dans le massif de la Clape, la logique saisonnière méditerranéenne crée, à l'automne et au printemps, des fenêtres où ces deux conditions peuvent se succéder assez rapidement pour que le cycle se boucle.
Certains stades du cycle ne durent que quelques jours et ne se lisent qu'à la loupe (×10 à ×50). Ce calendrier indique, mois par mois, ce qu'on peut chercher pour chacune des trois espèces : à l'œil nu, à la loupe ou au binoculaire, et où diriger le regard sur le terrain.
Sources : Page, 1997 · Peterson & Kott, 1974 · Van den Heede et al., 2003 · Tela Botanica, BDtfx · Prelli & Boudrie, 2002/2021 · observations terrain. Périodes indicatives pour le contexte méditerranéen de la Clape ; fenêtres à ajuster selon l'exposition et la pluviométrie annuelle.
Ce que ces quatre sections révèlent ensemble ne se lit pas dans chacune séparément. Les portraits décrivent des plantes. Le cadre environnemental décrit un massif. Les stratégies décrivent des mécanismes. Le cycle biologique décrit une contrainte. Mais c'est leur articulation qui dit quelque chose de plus précis : ces trois espèces occupent la même paroi calcaire parce qu'elles exploitent le même réservoir, l'épikarst, à des moments différents et par des voies différentes. Leur cohabitation n'est pas un hasard floristique. C'est la signature d'un régime hydrique particulier, celui d'un massif compartimenté, à recharge rapide et restitution lente. Quand ce régime se modifie, ce sont elles qui le signalent en premier.
Ce qu'il faut retenir
On est parti d'une question simple: comment des fougères survivaient dans un massif calcaire méditerranéen. Trois y sont parvenues, aucune de la même manière.
✦
Explorations AC et Ma Clape · Massif de la Clape, Aude
La prochaine fois que vous longerez une paroi calcaire dans la Clape et que vous apercevrez une touffe brunâtre et roulée sur elle-même dans une fissure, regardez de plus près. Elle n'est probablement pas morte.
Ces plantes sont là depuis longtemps, à des échelles de temps qui dépassent largement celle du climat actuel. Elles ont traversé des cycles climatiques bien plus sévères que les étés que nous connaissons. Ce qui les fragilise aujourd'hui n'est pas la sécheresse estivale, qu'elles tolèrent ou esquivent depuis des millions d'années. C'est la conjonction de deux modifications plus discrètes : les premières pluies d'automne arrivent plus tard, et la demande évaporatoire monte. Sur la même quantité d'eau tombée, l'eau utile diminue, et la fenêtre où la fécondation est possible se rétrécit. Le détail figure dans Les fenêtres de reproduction et l'encart Bilan hydrique .
Autres carnets du massif · Thématique Paysage & Nature
Flora Verticalis Part. I
Éloge de la verticalité dans le massif de la Clape
Sur les mêmes parois calcaires, d'autres plantes ont résolu autrement le problème de la verticalité. Les genévriers de la Clape, genévrier de Phénicie et genévrier cade, enracinés directement dans la roche: leur biogéographie, leur longévité, leur usage à travers les cultures méditerranéennes.
Lire ce carnet →La Centaurée de la Clape
Fleur rare, mémoire fragile
Espèce endémique dont l'aire mondiale se résume à six stations sur les falaises calcaires de ce massif. Elle pousse sur les mêmes parois que les fougères, dans les mêmes contraintes de sol squelettique et de sécheresse estivale. Un autre angle sur la même question: tenir sur la roche.
Lire ce carnet →Figuiers sauvages de la Clape
Arbres de la soif et du silence
Le caprifiguier de la Clape face à la même sécheresse, mais avec des stratégies radicalement différentes: un arbre, une guêpe, une interdépendance. La biologie de la pollinisation dans un milieu aride méditerranéen.
Lire ce carnet →Le Cers, maître des plaines et des lagunes
Météorologie et histoire d'un vent millénaire en Narbonnaise
L'ETP annuelle atteint 1241 mm sur la station Narbonne de référence, pour 635 mm de précipitations en moyenne (normale 1991-2020) : un déficit structurel de l'ordre de 600 mm avant même les épisodes de Cers. En rafale soutenue, ce vent sec renouvelle en continu la couche limite d'air humide autour des feuilles, et l'ETP réelle peut alors dépasser deux à trois fois sa valeur en conditions calmes. C'est le mécanisme derrière les 60 % d'évapotranspiration du bilan hydrique de la Clape.
Lire ce carnet →Tapez un terme ou cliquez une lettre pour afficher les définitions.
Qualifie la face inférieure de la fronde (le "dos"), opposée à la face adaxiale (le dessus). C'est sur cette face que se développent les sores chez les Polypodiacées.
Se dit d'une composante non vivante d'un écosystème, comme le climat, le sol, l'eau et la lumière
Se dit d'un organe qui se développe à un endroit inhabituel pour son type. Une racine adventive naît sur une tige, un rhizome ou une feuille, et non à partir d'une autre racine. Chez les fougères, les racines adventives partent directement du rhizome et assurent l'ancrage sur la roche.
Champignons mycorhiziens à arbuscules ( Arbuscular Mycorrhizal Fungi ). Ils forment une association symbiotique avec les racines des fougères leptosporangiées, étendant leur prospection phosphatée au-delà des pores inaccessibles aux radicelles et sécrétant des phosphatases qui libèrent le phosphore organique accumulé dans les fissures calcaires.
Mode de dispersion des spores assuré par le vent, favorisé par la légèreté des semences microscopiques
Jonction entre deux nervures qui se rejoignent pour former un réseau fermé de mailles. La nervation anastomosante s'oppose à la nervation libre, où les nervures se terminent en cul-de-sac sans se rejoindre. Chez A. ceterach , les anastomoses se forment près de la marge des pinnules, caractère propre au sous-genre Ceterach au sein des Aspleniaceae à nervation généralement libre. Présente également chez P. cambricum , absente chez P. vulgare . La fonction adaptative de cette structure n'a pas été étudiée directement chez ces espèces ; la redondance hydraulique est l'hypothèse la plus couramment avancée : un réseau fermé offre des chemins alternatifs de conduction si une veine est localement endommagée ou bloquée.
Forme extrême de tolérance à la dessiccation atteinte par certains organismes poïkilohydres Breuil, 1997 ; Proctor, 2009 . État dans lequel la déshydratation quasi totale ( RWC résiduelle 4 à 7 %, soit 93 à 96 % d'eau perdue chez le Cétérach Proctor, 2009 ) interrompt toutes les activités vitales mesurables, sans pour autant tuer l'organisme : la reprise est complète en quelques heures dès le retour de l'eau.
Arc de cellules à parois épaissies situé sur le sporange qui, en se rétractant par évaporation, provoque l'éjection des spores par effet de catapulte
Organe mâle du gamétophyte (prothalle) produisant les anthérozoïdes. Chez les fougères, elle se présente sous forme d'une petite capsule sphérique à paroi unicellulaire, libérant les gamètes mâles multiflagellés en présence d'eau liquide.
Gamète mâle mobile produit par le prothalle, portant de nombreux flagelles (multiflagellé), lui permettant de nager dans un film d'eau liquide
Forme minérale finale et hautement insoluble du phosphate de calcium, thermodynamiquement stable en milieu alcalin
Acide désoxyribonucléique. Molécule en double hélice portant l'information génétique de la cellule. Sa structure en nucléotides nécessite du phosphore pour former le squelette de la chaîne (groupements phosphate qui relient les sucres entre eux). C'est l'une des raisons pour lesquelles le phosphore est un nutriment irremplaçable pour toute cellule vivante.
Acide ribonucléique. Molécule issue de la transcription de l'ADN, impliquée dans la synthèse des protéines. Comme l'ADN, sa structure en nucléotides requiert du phosphore dans son squelette. Les fougères en produisent de grandes quantités lors de la réhydratation après une période de dessiccation, pour relancer la synthèse protéique.
Adénosine triphosphate. Molécule universelle de transfert d'énergie dans la cellule : chaque réaction biochimique qui consomme de l'énergie (synthèse de protéines, transport membranaire, mouvement) utilise l'ATP comme monnaie énergétique. Sa structure comporte trois groupements phosphate ; la libération du troisième libère l'énergie utilisable. Sans phosphore disponible, la cellule ne peut pas produire d'ATP et donc pas fonctionner.
Structure très ramifiée (en forme de petit arbre) formée par un champignon symbiotique à l'intérieur d'une cellule végétale, lieu d'échange des nutriments.
Organe femelle du gamétophyte (prothalle) contenant l'oosphère, cellule reproductrice femelle. Les anthérozoïdes doivent nager en film d'eau liquide jusqu'à l'archégone pour que la fécondation ait lieu. C'est ce point de passage qui fait de l'eau un prérequis absolu à la reproduction des fougères, même dans un milieu xérique.
Pellicule de micro-organismes (bactéries, cyanobactéries, algues) organisés en communauté sur une surface rocheuse, enchâssés dans une matrice polysaccharidique. Premier stade de colonisation de la roche nue: invisible à l'œil nu, reconnaissable collectivement aux traînées sombres ou grises sur les parois calcaires. Précède et conditionne l'installation des lichens.
Oligo-élément dont la disponibilité diminue en sol alcalin par adsorption spécifique sur les carbonates et les argiles
Premier phosphate de calcium précipité lors de la rétrogradation du phosphore en milieu calcaire (CaHPO₄·2H₂O). Encore partiellement soluble, elle est le stade intermédiaire entre le phosphore labile et l'hydroxyapatite quasi inerte. Sa formation est rapide dès pH > 7.
des plantes non vasculaires sans vraies racines, qui colonisent les substrats humides avant les fougères. Leur capacité à retenir l'eau de pluie et à ralentir l'évaporation du substrat conditionne l'installation des ptéridophytes sur la roche nue.
Fraction granulométrique fine des carbonates (< 20 𝜇𝑚) qui réagit rapidement avec la solution du sol
Se dit d'une plante qui a besoin d'un sol riche en calcium (calcaire) pour se développer. C'est le cas de la majorité des plantes du massif de la Clape.
Phénomène par lequel un liquide monte spontanément dans un espace étroit contre la gravité, sous l'effet de la tension superficielle et des forces d'adhésion aux parois. La hauteur de remontée est décrite par la loi de Jurin (1718) : h = 2γ cos θ / (ρgd), où d est la largeur de l'interstice. La relation est inverse : une fissure de 0,1 mm retient de l'eau jusqu'à ~14 cm au-dessus du front d'humectation ; une fissure de 1 mm ne la retient qu'à ~1,4 cm. Dans les fissures rocheuses étroites, ce mécanisme prolonge la disponibilité hydrique plusieurs jours après la pluie, indépendamment de tout apport extérieur.
Phénomène physique consistant en la formation brutale d'une bulle de gaz (vapeur) à l'intérieur des cellules de l'annulus soumises à une forte tension hydrique. C'est l'événement déclencheur qui libère l'énergie élastique pour éjecter les spores.
Plante colonisant spécifiquement les fentes et les fissures des parois rocheuses ou des vieux murs
Liaison chimique entre un cation et une molécule organique polyfonctionnelle qui le maintient en solution plutôt qu'à l'état précipité. Chez les fougères de la Clape, deux mécanismes distincts sont à l'œuvre: l'oxalate et le citrate se lient aux ions calcium des apatites calcaires pour libérer les phosphates piégés ; les coumarines et phénolates chélatent les ions fer (Fe³⁺ réduit en Fe²⁺) pour le rendre absorbable. Ces quatre composés ont leurs propres entrées dans ce glossaire.
Trouble physiologique se manifestant par le jaunissement des feuilles dû à l'insolubilité du fer en sol calcaire
Lichen dont le photobionte est une algue verte (le plus souvent du genre Trebouxia ), par opposition aux cyanolichens dont le photobionte est une cyanobactérie. C'est le type le plus répandu sur calcaire méditerranéen. Les chlorolichens ne fixent pas l'azote atmosphérique, mais ils peuvent réactiver leur photosynthèse à partir de la seule vapeur d'eau atmosphérique, sans eau liquide. Cette tolérance à la sécheresse en fait les pionniers lichéniques de la roche nue : sur calcaire, les crustacés Verrucaria et Aspicilia s'installent avant les cyanolichens.
Sel de l'acide citrique, excrété par les racines et rhizomes de fougères en situation de carence phosphatée. Le citrate chélate les ions calcium des apatites calcaires, libérant partiellement les phosphates piégés. Moins efficace que l'oxalate sur substrat calcaire pur, mais actif à des pH plus élevés.
Se dit d'une écaille dont les parois cellulaires forment un réseau sombre en grille, caractéristique du genre Asplenium
Interaction biologique où deux organismes luttent pour les mêmes ressources limitées, souvent remplacée par la facilitation en milieu hostile.
Se dit d'une feuille dont la texture est épaisse, tenace et résistante, rappelant le cuir. Une adaptation courante pour limiter la transpiration.
Composés phénoliques cycliques sécrétés par les racines en situation de carence ferrique. Avec les phénolates, ils forment la fraction chélatrice spécifique aux plantes non-graminées (Stratégie I): ils complexent le Fe³⁺ à la surface racinaire, facilitant sa réduction en Fe²⁺ par la ferréductase. Actifs à pH alcalin, ce qui les distingue des phytosidérophores des graminées.
Couche protectrice cireuse recouvrant l'épiderme des feuilles (frondes) pour imperméabiliser la plante et limiter la perte d'eau.
Bactérie photosynthétique. Certains genres ( Nostoc , Scytonema , Anabaena ) fixent l'azote atmosphérique grâce à des cellules spécialisées, les hétérocystes, où la nitrogenase opère à l'abri de l'oxygène. Sur calcaire, les cyanobactéries se rencontrent dans deux statuts : libres au sein du biofilm épilithique (premiers colonisateurs de la roche nue), ou en symbiose avec un champignon dans le thalle d'un cyanolichen.
Lichen dont le photobionte est une cyanobactérie ( Nostoc le plus souvent), par opposition aux chlorolichens dont le photobionte est une algue verte. Le thalle est typiquement gélatineux à l'état hydraté, sombre (vert olive à noir), foliacé, squamuleux ou crustacé selon le genre. Sur calcaire méditerranéen : Collema , Lathagrium , Enchylium , Scytinium . Contrairement aux chlorolichens, les cyanolichens fixent l'azote atmosphérique via le métabolisme de leur photobionte. Position successionnelle : ils s'installent en général plus tardivement que les chlorolichens crustacés ( Verrucaria , Aspicilia ), sur des surfaces déjà préparées par le biofilm et dans des microsites moyennement humides. La raison est physiologique : le photobionte cyanobactérien ne réactive sa photosynthèse qu'avec de l'eau liquide, là où l'algue verte des chlorolichens se réactive à partir de la seule vapeur d'eau atmosphérique. Le cyanolichen dépend donc de microsites qui retiennent l'eau liquide, absents de la roche nue.
Variante d'une espèce définie par un nombre chromosomique spécifique. Chez les fougères, une même espèce morphologique peut regrouper plusieurs cytotypes de niveaux de ploïdie différents (diploïde, tétraploïde) dont les exigences écologiques divergent nettement. Asplenium trichomanes en est l'exemple le mieux documenté en Europe: le cytotype diploïde (2n = 72, subsp. trichomanes ) est strictement calcifuge, tandis que le cytotype tétraploïde (2n = 144, subsp. quadrivalens ) tolère les substrats calcaires alcalins. La distinction est invisible à l'œil nu ; elle commande pourtant la répartition géographique des deux sous-espèces.
Processus d'ouverture spontanée d'un organe végétal clos (ici le sporange) pour libérer son contenu (les spores) à maturité.
Fissure tectonique traversant la roche calcaire, sans déplacement notable des deux blocs. Dans un massif karstique comme la Clape, les diaclases constituent les voies principales d'infiltration de l'eau et les premiers interstices colonisables par les plantes rupicoles, dont les rhizomes s'y ancrent et y prospectent l'humidité résiduelle.
Qualifie un organisme possédant deux jeux de chromosomes (2n). *Polypodium cambricum* est l'espèce diploïde (2n=74) ancestrale, distincte des espèces polyploïdes comme *P. vulgare* (tétraploïde).
Période géologique s'étendant de –419 à –359 Ma, appartenant à l'ère Paléozoïque. Le Dévonien moyen (~400 Ma) voit l'apparition des premières plantes vasculaires à feuilles et des premiers ancêtres des fougères actuelles. La morphologie générale des ptéridophytes est établie dès cette période et n'a pas fondamentalement changé depuis.
Qualifie des champignons dont les hyphes enveloppent les racines sans pénétrer dans les cellules, formant un manchon externe (manteau) et un réseau intercellulaire (réseau de Hartig). Ce mode d'association caractérise notamment Pisolithus arrhizus et Cenococcum geophilum documentés sur Cistus en substrat calcaire méditerranéen. À distinguer des mycorhizes à arbuscules (AMF), qui pénètrent dans les cellules racinaires et sont le type le plus commun chez les fougères.
Réseau souterrain de galeries, grottes et conduits creusés par la dissolution du calcaire par les eaux d'infiltration. Dans le massif de la Clape, une part de l'infiltration rejoint les conduits profonds de l'endokarst par les failles ; une autre reste piégée dans l'aquifère supérieur des calcaires urgoniens, bloquée par les marnes sous-jacentes. Ces deux circuits souterrains expliquent l'absence de cours d'eau permanents en surface Khaska et al., 2013 .
Qualifie un organisme qui vit et se développe sur la surface d'une roche, sans y pénétrer. Les cyanobactéries épilithiques adhèrent à la surface calcaire dans une gaine mucilagineuse ; leur activité prépare les premiers micromètres de la surface (micro-altération, rétention d'eau), créant la microrugosité nécessaire à l'ancrage des spores lichéniques. À distinguer de trois modes voisins: l'endolithique (l'organisme vit dans les pores naturels de la roche), l'euendolithique (il perfore activement la roche en y creusant des tunnels, comme Hyella ou Chroococcidiopsis ), et l'hypolithique (il vit sous les pierres). Sur les parois de la Clape, Nostoc (confirmé par observation directe) ainsi que, vraisemblablement, Gloeocapsa et Scytonema sont épilithiques: aucun de ces genres ne pénètre dans la roche.
Plante qui croît en se servant d'autres végétaux (comme les troncs d'arbres mousses) comme support, sans toutefois les parasiter pour se nourrir.

Zone altérée superficielle d'un massif calcaire,correspondant à la partie haute de l'UKU. Sa forte densité de micro-fissures lui confère une capacité de rétention capillaire: l'eau y reste piégée plusieurs jours à plusieurs semaines après une pluie, protégée de l'évaporation directe. Constitue le principal réservoir hydrique accessible aux plantes rupicoles, contrairement à l' endokarst profond, inaccessible aux racines.
Dormance estivale induite par la chaleur et le déficit hydrique; le rhizome reste viable et reprend son activité dès les premières pluies d’automne
Molécules organiques sécrétées par les racines pour modifier la chimie du sol et mobiliser les nutriments
Interaction écologique positive où une espèce "nurse" améliore les conditions pour permettre l'établissement d'une autre
Augmentation soudaine et massive de la proportion de spores de fougères dans les couches de sédiments fossiles. Ce phénomène se produit juste après une extinction massive, comme celle qui a causé la disparition des dinosaures.
Enzyme membranaire racinaire (FRO2 chez Arabidopsis , homologue supposé chez les fougères) qui réduit le Fe³⁺ ferrique insoluble en Fe²⁺ ferreux soluble, absorbable par les cellules. Son activité diminue significativement au-delà de pH 7, ce qui explique la persistance de la chlorose ferrique chez des plantes pourtant capables de réduction, sur substrats calcaires à pH 7,5–8,5 comme l'urgonien de la Clape.
Organe principal des fougères, jouant à la fois le rôle de feuille (photosynthèse) et de support pour la reproduction (portant les spores).
Phase haploïde (n) du cycle, issue de la germination de la spore et responsable de la production des gamètes
Groupe d'espèces (Mousses, Sédums, Fougères) s'associant localement pour exploiter les ressources et résister à la sécheresse.
Qualifie une cellule ou un organisme ne possédant qu'un seul jeu de chromosomes (n). Chez les fougères, les spores et le gamétophyte (prothalle) sont haploïdes. La fécondation réunit deux gamètes haploïdes pour former un zygote diploïde (2n), amorce du nouveau sporophyte.
Couche supérieure du sol créée par la décomposition de la matière organique, agissant comme une éponge pour retenir l'eau.
Qui repousse l'eau. La surface des feuilles de Capillaire est hydrophobe, empêchant l'eau de mouiller le limbe (effet lotus).
Ce dit d’une espèce qui exige une humidité élevée et durable pour se développer. S’oppose à xérophile (espèce adaptée à la sécheresse) et à mésophile (espèce des conditions intermédiaires). Les fougères sont généralement hygrophiles, ce qui explique leur rareté dans les milieux méditerranéens secs.
Filaments tubulaires microscopiques constituant le corps végétatif (mycélium) des champignons. Dans le contexte des associations mycorhiziennes, les hyphes explorent les micropores du substrat inaccessibles aux radicelles (< 2 µm de diamètre), multipliant par un facteur 10 à 100 la surface effective d'absorption de la plante. Ils mobilisent notamment le zinc, le bore et le manganèse adsorbés sur les surfaces calcaires.
Excroissance épidermique d'origine foliaire formant un tégument protecteur recouvrant le sore. Sa forme varie selon l'espèce: linéaire chez Asplenium trichomanes (liseré clair s'ouvrant vers la nervure à maturité), vestigiale et fonctionnellement remplacée par des paléas chez Asplenium ceterach, absente chez Polypodium cambricum (sores nus).
Fraction des précipitations qui atteint effectivement la zone saturée de l'aquifère (nappe), après déduction de l'évapotranspiration, du ruissellement et de l'eau retenue dans la zone non saturée (épikarst). Sur un karst méditerranéen de basse altitude, elle représente environ 35 % des précipitations annuelles en moyenne ; les épisodes méditerranéens intenses peuvent la faire varier brutalement. Sur la Clape, ce terme s'applique avec réserve : la structure tripartite du massif (calcaires urgoniens / marnes / calcaires profonds) compartimente l'infiltration entre un aquifère supérieur à résurgences temporaires et un aquifère profond scellé, dont la recharge active n'est pas quantifiée Khaska et al., 2013 .
Paysage et système hydrologique résultant de la dissolution chimique d'une roche carbonatée (calcaire, dolomie) par les eaux chargées en CO₂. Ses caractéristiques: absence de réseau hydrographique de surface, drainage souterrain quasi total, intercepté sur la Clape par une unité marneuse qui compartimente l'infiltration entre un aquifère supérieur à résurgences temporaires et un réseau de galeries profondes Khaska et al., 2013 , et morphologie de surface sculptée en lapiaz, dolines et avens. Le massif de la Clape est un karst urgonien méditerranéen.
Qualifie la fraction d'un nutriment (notamment le phosphore) facilement mobilisable dans la solution du sol, par opposition aux formes fixes ou précipitées. Le phosphore organique libéré par les mousses est plus labile que les apatites de la roche brute: c'est sur cette fraction que les fougères puisent en priorité.
Surface calcaire creusée de stries, rigoles et fissures par la dissolution lente de la roche sous l'action des eaux de ruissellement chargées en CO₂. Dans le massif de la Clape, les fissures du lapiaz constituent les seules micro-poches de sol où lichens, mousses et fougères peuvent s'installer.
Groupe de fougères possédant des sporanges à paroi fine issus d'une seule cellule initiale
Organisme symbiotique formé par l'association d'un champignon et d'un partenaire photosynthétique : une algue verte chez les chlorolichens (la grande majorité des espèces de la Clape), ou une cyanobactérie chez les cyanolichens ( Collema , Lathagrium , Scytinium ). Pionnier rupicole capable de préparer le substrat pour l'installation des mousses puis des plantes vasculaires.
Lignée de plantes vasculaires (Lycopodiopsida) divergeant des Euphyllophytes au Dévonien moyen (~400 Ma). Comprend les lycopodes, sélaginelles et isoètes actuels. Dominant dans les forêts du Carbonifère sous forme arborescente, les Lycophytes sont aujourd'hui marginaux. Aucun représentant n'est présent dans la Clape.
Biopolymère rigide et hydrophobe imprégnant les parois cellulaires. Sa couleur naturelle jaune/ambrée est responsable de la teinte dorée brillante de l'annulus à maturité.
Partie large, aplatie et généralement verte d'une feuille ou d'une fronde, siège principal de la photosynthèse.
Sol squelettique formé directement sur la roche-mère, avec un horizon organique quasi inexistant (épaisseur inférieure à 10 cm). Dans le massif de la Clape, les lithosols des fissures de lapiaz sont soumis à des fluctuations hydriques et thermiques extrêmes. Ils constituent pourtant le seul substrat disponible pour les chasmophytes.
Du grec lithos (pierre) et tribein (broyer). Qualifie une plante ou une préparation réputée dissoudre ou fragmenter les calculs urinaires et rénaux. Terme de la pharmacopée ancienne, aujourd'hui abandonné en médecine. Asplenium ceterach était réputé lithontripique, ce qui lui valut longtemps le nom d'espèce officinarum (de l'officine). Aucune efficacité thérapeutique n'a été démontrée par des études cliniques contrôlées.
Division cellulaire réductrice qui transforme une cellule diploïde (2n) en quatre cellules haploïdes (n). Chez les fougères, elle se produit dans les sporanges: les cellules mères diploïdes du sporophyte y subissent la méiose pour produire les spores haploïdes. C'est l'étape qui rompt la diploïdie et amorce la génération gamétophytique. Sans diploïdie en point de départ, pas de méiose possible: le cycle ne peut pas s'accomplir.
Ère géologique s'étendant de –252 à –66 Ma, subdivisée en Trias, Jurassique et Crétacé. Le Mésozoïque est la période de radiation majeure des fougères modernes : les familles actuelles des Polypodiales, dont les trois espèces de ce carnet, s'y diversifient et s'y établissent sur les substrats rocheux. L'aquifère calcaire de la Clape est constitué de calcaires crétacés formés au cours de cette ère ( Khaska et al., 2013 ).
Zone de tissu végétal dont les cellules restent indifférenciées et conservent la capacité de se diviser activement. Chez les fougères, le méristème apical du rhizome produit en continu de nouvelles frondes par l'allongement du rhizome. Les méristèmes sont les premiers tissus à souffrir d'une carence en phosphore, car la synthèse d'ADN et d'ARN nécessaire à chaque division cellulaire en requiert de grandes quantités.
Conditions climatiques spécifiques à une zone très restreinte (fissure, sous une pierre) qui permettent la survie d'espèces sensibles.
Cavités du substrat dont le diamètre est inférieur à 30 µm, inaccessibles aux radicelles (diamètre minimal 50–150 µm) mais explorables par les hyphes fongiques (2–7 µm). Dans les fissures calcaires de la Clape, les micropores concentrent le phosphore organique labile issu de la décomposition des mousses et des lichens. C'est précisément cette fraction que les champignons mycorhiziens arbusculaires (AMF) peuvent mobiliser, là où les racines des fougères ne peuvent pas atteindre.
Association symbiotique entre champignons et racines améliorant l'exploration du sol pour le phosphore
Sel de l'acide oxalique, principal exsudat racinaire mobilisateur de phosphore en milieu calcaire. L'oxalate chélate le calcium des apatites (Ca₁₀(PO₄)₆(OH)₂), libérant des ions phosphate en solution. Sécrété par les rhizomes d' Asplenium et de Polypodium en situation de carence, il crée une zone rhizosphérique chimiquement distincte de quelques millimètres autour de l'organe souterrain.
Écaille épidermique membraneuse, souvent de couleur rousse ou brune, recouvrant le rhizome et la face inférieure des frondes (notamment chez les xérophytes) pour limiter l'évaporation.
Ensemble des processus conduisant à la formation et à l'évolution des sols
Filament cellulaire stérile, parfois ramifié, intercalé entre les sporanges sur le sore de certaines fougères. Chez les Polypodium d'Europe occidentale, le degré de ramification sépare les trois espèces voisines: paraphyses nombreuses et ramifiées chez P. cambricum , peu ramifiées chez P. interjectum , absentes chez P. vulgare . Visible à la loupe binoculaire sur sore mature. Leur rôle fonctionnel n'est pas établi sur P. cambricum ; chez le congénère P. virginianum , il a été proposé une fonction de protection des sporanges en développement contre la dessiccation Peterson & Kott, 1974 .
Se dit d'une feuille dont le limbe est divisé en lobes profonds, mais dont les divisions n'atteignent pas complètement la nervure centrale (rachis).
Se dit d'une feuille dont le limbe est profondément découpé en segments (pennes) atteignant la nervure centrale.
Division primaire du limbe d'une fronde de fougère (équivalent à la foliole chez les plantes à fleurs). Les sores sont disposés sur les pennes.
Mesure de l'acidité ou de l'alcalinité régissant les équilibres de solubilité dans la solution du sol
Sels ou esters d'acides phénoliques sécrétés par les racines en condition de carence ferrique. Associés aux coumarines dans la Stratégie I de mobilisation du fer chez les plantes non-graminées: ils complexent le Fe³⁺ à la surface racinaire, favorisant sa réduction par la ferréductase. Leur efficacité se maintient en milieu alcalin, là où les phytosidérophores des graminées sont moins actifs.
Étude de l'apparition d'événements périodiques de la vie végétale (comme le développement des frondes ou la maturation des spores) en relation avec les variations saisonnières du climat.
Membrane externe de la spore, déposée tardivement par les cellules du tapetum sporangial. Souvent ornée (plis, verrues, crêtes), elle porte les caractères morphologiques utilisés pour identifier les espèces au microscope. Chez les Aspleniaceae, le périspore est plissé ; chez Polypodium, il est verruqueux et jaune doré.
Dommage causé à l'appareil photosynthétique par un excès de lumière, souvent aggravé par la sécheresse
Voie photosynthétique la plus répandue, celle de la majorité des plantes dont les fougères. Les stomates s'ouvrent le jour pour capter le CO₂, fixé directement en une molécule à trois atomes de carbone (d'où le nom). Efficace en conditions tempérées, mais coûteuse en eau sous climat chaud et sec : l'ouverture diurne des stomates coïncide avec le pic de demande évaporative. C'est la limite que contourne la photosynthèse CAM des plantes succulentes.
Métabolisme acide crassulacéen ( Crassulacean Acid Metabolism ), voie photosynthétique des plantes succulentes (sédums, cactées, agaves). Les stomates s'ouvrent la nuit, quand l'air est plus frais et humide, pour fixer le CO₂ sous forme d'acide organique stocké jusqu'au jour. Cette inversion réduit fortement les pertes d'eau par rapport à la photosynthèse C3 , au prix d'une croissance plus lente. Sur le massif, c'est la stratégie des Sedum qui cohabitent avec les fougères dans les fissures.
Molécules chélatrices du fer (principalement la mugineïque acide et ses dérivés) sécrétées par les graminées et certaines monocotylédones pour mobiliser le Fe³⁺ en milieu alcalin (Stratégie II). Absents chez les fougères et la plupart des dicotylédones, qui utilisent à la place la voie réductive (ferréductase + coumarines/phénolates, Stratégie I). Le contraste entre les deux stratégies est utile pour comprendre pourquoi la chlorose ferrique frappe différemment les espèces sur le même substrat calcaire.
Relatif à la phylogénie, c'est-à-dire à l'histoire évolutive et aux relations de parenté entre organismes. Une classification phylogénétique regroupe les espèces selon leur ascendance commune, indépendamment de leur ressemblance morphologique. Dans ce carnet, les relations phylogénétiques entre Asplenium ceterach et A. trichomanes expliquent pourquoi deux espèces cohabitant dans la même fissure n'hybrident pas ( Schneider et al., 2004 ).
Molécules constituant la double couche lipidique de toutes les membranes cellulaires. Chaque phospholipide comporte une tête polaire hydrophile (contenant un groupement phosphate) et deux queues lipidiques hydrophobes. Leur organisation en bicouche crée la barrière semi-perméable qui sépare l'intérieur de la cellule du milieu extérieur et contrôle les échanges. La proportion de phospholipides insaturés dans les membranes détermine leur fluidité à basse température, ce qui conditionne la résistance au gel des fougères en hiver.
Espèce végétale capable de coloniser en premier un milieu vierge ou difficile comme la roche calcaire nue.
État dans lequel un organisme ne régule pas activement sa teneur en eau, contrairement aux plantes terrestres typiques qui contrôlent leurs échanges par la cuticule et les stomates. Une plante poïkilohydre se dessèche quand l'air est sec et se réhydrate quand l'humidité revient. Toutes les fougères sont poïkilohydres à des degrés divers ; seules les plus tolérantes, comme le Cétérach, atteignent l' anhydrobiose , état dans lequel la déshydratation extrême interrompt toutes les activités vitales mesurables (respiration, photosynthèse, croissance), sans pour autant tuer l'organisme : la reprise est complète dès que l'eau revient Proctor & Tuba, 2002 .
État chromosomique caractérisé par la possession de plus de deux jeux complets de chromosomes
Ordre de fougères leptosporangiées apparu au Jurassique (180–160 Ma selon Schuettpelz & Pryer 2009 ; des estimations plus récentes, Du et al. 2021 et Nitta et al. 2022, repoussent l'origine jusqu'au Trias, sans consensus établi), regroupant la grande majorité des fougères modernes. Elle comprend les familles Polypodiaceae ( Polypodium ) et Aspleniaceae ( Asplenium ). Leur radiation évolutive, coïncidant avec l'expansion des forêts d'angiospermes au Crétacé, a produit plus de 10 000 espèces actuelles, dont les trois espèces de la Clape.
Protéines protectrices s'accumulant lors de la dessiccation pour stabiliser les membranes et les enzymes
Accumulation organo-minérale initiale dans les anfractuosités d'une roche nue: quelques millimètres à quelques centimètres de matière décomposée (débris végétaux, particules fines, exsudats microbiens), sans horizons différenciés. Précède la formation d'un sol au sens pédologique. Sur le calcaire de la Clape, il est constitué principalement de la décomposition des lichens, des mousses et des sédums. Seuil minimal pour la germination des spores de fougères rupicoles.
Gamétophyte des fougères, prenant souvent la forme d'un petit cœur vert fixé au substrat
Embranchement végétal regroupant les plantes vasculaires ne produisant pas de graines (fougères, prêles), se reproduisant par des spores.
Axe central de la fronde, prolongeant le stipe (pétiole) et portant le limbe ou les pennes.
Processus de passage d'un nutriment de l'état soluble à une forme solide cristalline indisponible
Source où les eaux d'infiltration souterraine rejoignent la surface. Dans la Clape, les résurgences temporaires émergent à la base des calcaires urgoniens supérieurs, à leur contact avec les marnes sous-jacentes qui bloquent la percolation vers le bas ; elles sont conditionnées aux épisodes pluvieux et trahissent le transit dans l'aquifère supérieur. L'aquifère profond (LKU), scellé par les dépôts tertiaires environnants, ne produit pas de résurgences accessibles en surface Khaska et al., 2013 . Les fougères rupicoles n'en dépendent pas directement.
Tige souterraine ou rampante assurant l'ancrage et le stockage des réserves
Zone de sol située à proximité immédiate des racines et influencée par l'activité biologique végétale
Qualifie un organisme vivant sur ou dans les rochers (du latin rupes , le rocher). Une plante rupestre occupe les parois, fissures ou éboulis rocheux comme habitat principal. Synonyme partiel de saxicole, mais d'usage plus large: une mousse rupestre peut coloniser aussi bien une falaise verticale qu'un bloc isolé en garrigue.
Teneur en eau relative, de l'anglais relative water content . Rapport entre l'eau présente dans un tissu et l'eau maximale qu'il peut contenir à pleine turgescence : RWC = (masse fraîche − masse sèche) / (masse turgide − masse sèche) × 100. À 100 %, le tissu est saturé ; à 0 %, totalement sec. La plupart des tissus végétaux meurent dès qu'une RWC d'environ 30 % est atteinte ; les fougères tolérantes à la dessiccation comme le Cétérach descendent à 4-7 % et reprennent leur activité après réhydratation.
Se dit d'une plante qui vit fixée sur les rochers (du latin *saxum*, rocher).
Théorie écologique stipulant que l'entraide (facilitation) devient dominante sur la compétition lorsque le milieu est très stressant.
Amas de sporanges groupés sur la face inférieure de la fronde
Lignée de plantes vasculaires (Equisetopsida) divergeant des autres Euphyllophytes au Dévonien. Arborescente et abondante dans les forêts du Carbonifère sous la forme du genre fossile Calamites (jusqu'à 20 m de haut), elle est réduite aujourd'hui à la prêle ( Equisetum , seul genre survivant). Aucun représentant n'est présent dans la Clape.
Capsule microscopique au sein de laquelle sont produites les spores
Cellule reproductrice haploïde (n) microscopique, assurant la dispersion de l'espèce et donnant naissance au prothalle après germination.
Phase diploïde (2n) et feuillée du cycle de vie des fougères
Polymère biologique extrêmement résistant qui constitue la couche externe des spores, les protégeant de la dessiccation et des rayons UV.
Chez les fougères, correspond au pétiole de la feuille (la "queue" de la fronde avant les premières feuilles).
Mode d'acquisition du fer utilisé par les plantes non-graminées (dicotylédones, fougères). La plante acidifie la rhizosphère par pompage de H⁺, puis une ferréductase membranaire convertit le Fe³⁺ insoluble en Fe²⁺ soluble, absorbable par les cellules racinaires. Des exsudats chélateurs (coumarines, phénolates) accompagnent la réduction. Actif à pH alcalin, contrairement à la Stratégie II des graminées, qui repose sur des phytosidérophores sélectifs pour Fe³⁺. Source : Römheld & Marschner, 1986.
Association durable et intime entre deux organismes d'espèces différentes, souvent bénéfique aux deux parties (mutualisme).
Sol résiduel rouge-brique caractéristique des karsts calcaires méditerranéens, formé par accumulation des oxydes de fer et d'argile insoluble laissés après dissolution du calcaire. Dans les fissures du lapiaz de la Clape, la terra rossa est chimiquement riche en fer total, mais le pH alcalin maintient ce fer sous forme d'oxydes inaccessibles aux plantes.
Qualifie un organisme possédant quatre jeux complets de chromosomes (4n), contre deux chez le diploïde. Chez Asplenium trichomanes , la sous-espèce quadrivalens est tétraploïde (2n = 144) et tolère les substrats calcaires alcalins, là où le cytotype diploïde (2n = 72) est strictement calcifuge. La polyploïdie est un mécanisme fréquent d'adaptation écologique chez les fougères.
Corps végétatif des organismes non différenciés en racine, tige et feuilles (algues, lichens, hépatiques). Chez les lichens, le thalle est la structure principale visible: crustacé (soudé à la roche, indécollable), foliacé (lobé, partiellement détachable) ou fruticuleux (dressé). Sa forme détermine le rôle du lichen dans la succession écologique.
Se dit d'un mouvement végétal déclenché par une variation de température, indépendant de la direction du stimulus (par opposition au tropisme, qui suit une direction). Chez Asplenium trichomanes, l'enroulement des pinnules par forte chaleur estivale réduit la surface exposée au rayonnement solaire et limite les pertes par transpiration. La réponse est réversible: les pinnules se déploient à la fraîche.
Faciès de calcaire blanc à gris du Crétacé inférieur (Aptien, –125 à –113 Ma), très dur, à grains fins, riche en rudistes et autres organismes récifaux fossiles. C'est la roche dominante du massif de la Clape. Sa dureté élevée et son fort taux de CaCO₃ (souvent > 95 %) en font le substrat type du verrou nutritif décrit dans ce chapitre.
Enroulement caractéristique en "crosse" des jeunes frondes en cours de développement
Caractère d'un milieu structurellement déficient en eau disponible. Un habitat xérique cumule faibles précipitations, évaporation élevée et substrat à faible rétention hydrique. Dans le massif de la Clape, la xéricité estivale est un facteur de sélection direct: les espèces sans adaptation au déficit hydrique n'y persistent pas.
Se dit d'une plante adaptée aux milieux secs et arides (du grec *xeros*, sec).
Bibliographie
Les références ci-dessous correspondent aux sources citées dans le texte, les encarts et le glossaire. Elles sont organisées par thème. Les données pluviométriques locales proviennent d'une station privée (Narbonne-ville) croisée avec les normales Météo-France (station 11262005 · 110 m alt.).
✦ ✦ ✦
·
·































{kind=link}